Carlos Eduardo Maia Gomes. Fatores genéticos e étnicos relacionados à evolução da infecção por Leishmania infantum
|
|
|
- Isaac Teves Ávila
- 8 Há anos
- Visualizações:
Transcrição
1 Carlos Eduardo Maia Gomes Fatores genéticos e étnicos relacionados à evolução da infecção por Leishmania infantum Orientadora: Profª Drª Selma Maria Bezerra Jerônimo Natal/RN 2011 I

2 Carlos Eduardo Maia Gomes Fatores genéticos e étnicos relacionados à evolução da infecção por Leishmania infantum Tese apresentada à Universidade Federal do Rio Grande do Norte como requisito parcial para a obtenção do título de Doutor em Ciências da Saúde Orientadora: Profª Drª Selma Maria Bezerra Jerônimo Natal/RN 2011 II

3 Fatores genéticos e étnicos relacionados à evolução da infecção por Leishmania infantum Tese apresentada à Universidade Federal do Rio Grande do Norte como requisito parcial para a obtenção do título de Doutor em Ciências da Saúde Orientadora: Profª Drª Selma Maria Bezerra Jerônimo IV

4 Catalogação da Publicação na Fonte. UFRN / Biblioteca Setorial do Centro de Biociências Gomes, Carlos Eduardo Maia. Fatores genéticos e étnicos relacionados à evolução da infecção por Leishmania infantum / Carlos Eduardo Maia Gomes. Natal, RN, f. : Il. Orientadora: Profa. Dra. Selma Maria Bezerra Jerônimo. Tese (Doutorado) Universidade Federal do Rio Grande do Norte. Centro de Biociências. 1. Leishmaniose visceral Tese 2. Etnia Tese. 3. Genética Tese. I. Jerônimo, Selma Maria Bezerra. II. Universidade Federal do Rio Grande do Norte. III. Título. RN/UF/BSE-CB CDU III

5 CARLOS EDUARDO MAIA GOMES Fatores genéticos e étnicos relacionados à evolução da infecção por Leishmania chagasi Tese apresentada ao Programa de Pós-Graduação em Ciências da Saúde da Universidade Federal do Rio Grande do Norte como requisito parcial para obtenção do título de Doutor em Ciências da Saúde. Aprovado em: 16/12/2011 BANCA EXAMINADORA Profª Dra. Selma Maria Bezerra Jerônimo Departamento de Bioquímica - UFRN Orientadora Prof. Dr. Wagner Franco Molina Departamento de Biologia Celular e Genética UFRN 1o Examinador Interno Profª. Dra. Joanlise Marco Leon Andrade Departamento de Estatística UFRN 2ª Examinadora Intena Profª. Dra. Lea Cristina de Carvalho Castellucci Serviço de Imunologia Hospital Universitário Prof Edgard Santos UFBA 1ª Examinadora Externa Profª. Drª. Maria Goretti de Freire Carvalho Departamento de Patologia UNP 2o Examinador 3

6 Agradecimentos À Profª. Dra. Selma Maria Bezerra Jerônimo, pela orientação, apoio e confiança durante todo o curso de doutorado. Obrigado por aturar meu temperamento. À Dra. Jenefer Blackwell por me acolher em seu laboratório e à Dra. Sarra Jamieson por me acolher em sua cidade e pelos muitos ensinamentos no curtíssimo período em que trabalhamos juntos. Ao Dr. Edgar Carvalho apoio científico na realização deste trabalho. Aos Drs. Mary E. Wilson (Universidade de Iowa), Richard D. Pearson (Universidade da Virginia) e Priya Duggal (Johns Hopkins University) pelo apoio científico na realização dos estudos clínicos e genéticos. A Dra. Priya Duggal realizou a análise de linkage considerada neste estudo. Aos membros componentes da Banca de Defesa, Profª Drª Lea Castelucci, Profª Drª Goretti Carvalho, Profª Drª Joanlise Andrade e Prof Dr Wagner Molina pela análise e contribuições muito importantes para este trabalho. Aos professores membros da Banca de Qualificação Profº Dr. Wagner Franco Molina e Profª Dra. Lara de Melo Barbosa pelas avaliações criteriosas, pertinentes e muito valiosas para este trabalho. Aos meus pais e minha irmã, pelo apoio incondicional e estímulo. 1
 e Priya Duggal (Johns Hopkins University) pelo apoio científico na realização dos estudos clínicos e genéticos. A Dra.")
7 A meus tios, Sebastião e Francisco Maia, meus guias, por me colocarem no caminho certo. Aos amigos e colegas do Departamento de Biofísica e Farmacologia pelo ambiente cordial e tranquilo, que muito contribui para o bom andamento dos trabalhos presentes e futuros. Aos meus caríssimos velhos e novos amigos Alexandre Queiroz, Aulus Barbosa, Bruno Maggi, Frederico Lemos, Leonardo Capistrano, Ivan Rui, Sérgio Araújo, Patrícia Mesquita, Thiago Sanna e Jaime Groff pelo incentivo, pelo carinho e por me suportar. Aos colegas do Laboratório de imunogenética pela convivência relativamente pacífica. Aos queridos alunos do Laboratório de Biofísica Molecular pelo apoio e carinho. A todos os professores e funcionários do Departamento de Bioquímica, por me receberem tão bem no Departamento e por tudo o que aprendi. Aos Institutos Nacionais de Saúde dos EUA pelo financiamento do projeto de pesquisa que executei e me que permitiu a obtenção deste título de doutorado. 2

8 Sumário RESUMO INTRODUÇÃO OBJETIVOS REVISÃO DA LITERATURA ASPECTOS GERAIS RESPOSTA IMUNE À LEISHMANIA VARIAÇÃO GENÉTICA NA RESPOSTA IMUNE: ETNIA E SUSCEPTIBILIDADE A LEISHMANIOSE VISCERAL BRASIL E SUDÃO MATERIAL E MÉTODOS VARREDURA DE GENOMA ÁREA DE ESTUDO EXAMES FÍSICOS E ESTUDOS LABORATORIAIS FAMÍLIAS E AMBIENTE DEFINIÇÕES DOS FENÓTIPOS GENOTIPAGEM DOS MARCADORES MICROSSATÉLITE: CONTROLE DE QUALIDADE DOS DADOS: ANÁLISES ESTATÍSTICAS DA VARREDURA DE GENOMA: CONSIDERAÇÕES ÉTICAS ESTRUTURA ÉTNICA DE POPULAÇÕES DO NORTE E NORDESTE DO BRASIL E REANÁLISE DOS MARCADORES DA VARREDURA DE GENOMA DA POPULAÇÃO DE NATAL RN ÁREAS DE ESTUDO PARTICIPANTES GENOTIPAGEM DE MARCADORES DO CROMOSSOMO Y ANÁLISE DE ETNIA COMPARAÇÃO DAS FREQUÊNCIAS DE GRUPOS ÉTNICOS ENTRE DIFERENTES FENÓTIPOS ASSOCIADOS À INFECÇÃO POR L. INFANTUM ANÁLISE DE ASSOCIAÇÃO DOS DADOS DA VARREDURA DE GENOMA EM FUNÇÃO DOS ESTRATOS ÉTNICOS RESULTADOS E DISCUSSÃO VARREDURA DE GENOMA CONSIDERANDO A DOENÇA LEISHMANIOSE VISCERAL ESTRUTURA ÉTNICA: VARIAÇÃO GENÉTICA ENTRE AS POPULAÇÕES COMPARAÇÃO DAS FREQUÊNCIAS DE GRUPOS ÉTNICOS ENTRE DIFERENTES FENÓTIPOS ASSOCIADOS À INFECÇÃO POR L. INFANTUM ANÁLISE DE ASSOCIAÇÃO DOS DADOS DE VARREDURA DO GENOMA COM FENÓTIPOS RELACIONADOS À INFECÇÃO POR L. INFANTUM EM FUNÇÃO DA ORIGEM ÉTNICA DISCUSSÃO GERAL: CONCLUSÕES REFERÊNCIAS BIBLIOGRÁFICAS ANEXO

9 Lista de Figuras Figura Descrição Página Figura 1 Mapa do Brasil indicando os estados de onde foram 31 arrolados participantes da pesquisa Figura 2 Relações filogenéticas entre os marcadores étnicos de 35 cromossomo Y utilizados no presente estudo Figura 3 Sumário dos resultados da varredura de genoma 43 considerando os fenótipos leishmaniose visceral e infecção assintomática por Leishmania (resposta DTH) Figura 4 Mapa do Brasil indicando as freqüências de cada grupo étnico nos locais estudados 50 5
 Figura 4 Mapa do Brasil indicando as freqüências de cada grupo étnico")
10 Lista de Tabelas Tabela Descrição Página Tabela 1 Origem das famílias participantes do estudo com a 34 respectiva indicação dos tipos de doenças relacionadas ao recrutamento inicial Tabela 2 Número de famílias e participantes do estudo com os seus 39 respectivos fenótipos resultantes da exposição à Leishmania infantum. As famílias foram recrutadas devido a história de LV, certificada por registros hospitalares ou por residirem próxima a um caso de LV.. Os tipos de fenótipos são explicados no texto.. Tabela 3 Características das 1254 pessoas oriudas do estudo de 41 linkage considerando exposição à Leishmania infantum. Tabela 4 Principais resultados da análise de ligação em todo o 45 genoma da resposta de Montenegro (DTH), com as análises correspondentes para LV nas mesmas regiões Tabela 5 Freqüências relativas (%) dos haplogrupos de cromossomo 51 Y encontradas nas áreas estudadas Tabela 6 Freqüências absolutas e relativas dos haplogrupos e 52 haplótipos europeus encontrados nas famílias estudadas Tabela 7 Análise de variância molecular dentro e entre as populações 55 estudadas como um todo, e com relação aos haplogrupos e haplótipos africanos Tabela 8 Valores de distância genética (F st ) entre as populações 56 estudadas Tabela 9 Análise de significância de associação entre marcadores de 60 microssatélite e desfecho da infecção em indivíduos agrupados por origem étnica Tabela 10 Freqüências alélicas relativas do marcador D9S

11 Lista de quadros Quadro Descrição Página Quadro 1 Painéis de marcadores do cromossomo Y utilizados 35 Quadro 2 Marcadores genéticos utilizados na análise de 39 associação com desfechos da infecção por L. infantum em função da origem patrilinear dos participantes da pesquisa Quadro 3 Picos da varredura de genoma considerando os fenótipos LV e DTH 46 7

12 Lista de abreviações LV Leishmaniose visceral DTH Delayed-Type Hipersensitivity AC Anticorpo ELISA Enzyme-Linked Immunosorbent Assay PCR - Polymerase Chain Reaction PCR Th Célula T helper IL Interleucina NK Natural Killer INF- - Interferon- TNF- - Fator de necrose tumoral NFAT - Nuclear Factor of Activated T cells HIV Human Immunodeficiency Vírus TGF- - Transforming Growth Factor- SNP - Single Nucleotide Polymorphisms DNA Desoxiribonucleic Acid AMOVA Analysis of molecular variance NO- Óxido nítrico NCBI National Center for Biotechnology Information 8

13 Resumo: A população brasileira é composta por três subpopulações originais: ameríndios autóctones, exploradores europeus e escravos africanos. No Nordeste do Brasil há áreas endêmicas para diversas doenças causadas por microorganismos intracelulares como Leishmania spp e Mycobacterium. A susceptibilidade ou resistência a essas infecções parece em parte ser influenciada geneticamente. Estudos realizados na área endêmica para leishmaniose visceral (LV) no Rio Grande do Norte demonstram a existência de agregação familiar de infecção por Leishmania. Entre tais estudos está uma varredura de genoma a qual identificou três regiões cromossômicas associadas a evoluções distintas frente à infecção por Leishmania infantum. No presente estudo nós determinamos as proporções de ancestralidade paterna de pessoas residentes na área endêmica para LV no Rio Grande do Norte, Bahia e Belém através de marcadores genéticos do cromossomo Y. Nos indivíduos oriundos do RN as frequências alélicas de marcadores moleculares extratificados por origem étnica foram analisados buscando associação com doença e infecção assintomática. Cerca de 75% da contribuição paterna das populações estudadas é européia, com 20% de contribuição africana e o restante ameríndia. Foi detectada a presença de ancestralidade judia. A maior parte da variação genética observada devese ao componente africano. A análise de relação entre origem étnica exclusivamente paterna e desfecho da infecção indicou que não há associação entre essas variáveis. A análise de associação de marcadores microssatélites mostrou a associação de um marcador (D9S925) com leishmaniose visceral. Esses resultados tomados em conjunto parecem indicar que na população da área endêmica no RN a etnia influencia na freqüência alélica de genes que podem interferir no o desfecho da infecção por L. infantum. 9
 no Rio Grande do Norte demonstram a existência de agregação familiar de infecção por Leishmania.")
14 1. INTRODUÇÃO O termo leishmaniose refere-se a um espectro de doenças causadas por protozoários do gênero Leishmania. As doenças causadas por diferentes espécies de Leishmania variam e, embora exista sobreposição, cada espécie pode causar uma ou mais doenças. A leishmaniose visceral (LV), que normalmente é causada por Leishmania donovani ou Leishmania infantum, pode ser fatal mesmo com tratamento. Entretanto, mesmo entre pessoas infectadas com a mesma espécie de Leishmania há uma variabilidade considerável no desfecho clínico. Fatores ambientais influenciam, mas não explicam totalmente essas diferenças no desfecho da infecção. Essas observações levaram pesquisadores a examinar fatores inerentes ao hospedeiro que poderiam influenciar a expressão clínica da infecção. A leishmaniose visceral era uma doença mais comum nas áreas rurais do Brasil, mas desde os anos 80 a migração humana das áreas rurais para as urbanas resultou em epidemias nas principais cidades da região Nordeste (1-4). A existência dessas regiões onde há alto grau de exposição tem permitido estudos de fatores genéticos humanos que colocam as pessoas sob risco de desenvolver a doença. O desfecho assintomático da infecção é particularmente importante porque as pessoas podem desenvolver uma resposta imune tipo I ao parasito e parecem estar protegidos de desenvolver formas sintomáticas enquanto são imunocompetentes, mesmo com possível contínua reexposição a Leishmania. No entanto, a demonstração que a Leishmania é um microorganismo que pode se comportar de forma oportunista, resulta em preocupação crescente devido ao aumento de pessoas em risco de infecção 10

15 em áreas endêmicas da América Latina e, portanto, com possibilidadade de manisfestar a LV, com resposta dimunuída ao tratamento específico. Objetivos 1. Avaliar se há regiões no genoma humano associados a evoluções clínicas distintas em indivíduos infectados por L. infantum no Rio Grande do Norte; 2. Avaliar a ancestralidade paterna de indivíduos residentes em áreas endêmicas para infecções por parasitos intracelulares no Norte e Nordeste do Brasil; 3. Testar associação entre marcadores genéticos e o desfecho da infecção por L. infantum estratificando por origem étnica. 11

16 2. REVISÃO DA LITERATURA 2.1 Aspectos Gerais A Leishmaniose Visceral (LV) é uma doença complexa causada por protozoários do gênero Leishmania (Kinetoplastida; Trypanosomatidae) que são parasitos intracelulares obrigatórios de macrófagos (5). A infecção por este protozoário pode resultar em um quadro assintomático, detectado apenas por uma reação positiva à presença de antígenos do parasito (teste de Montenegro), ou em uma doença disseminada, caracterizada por hepatoesplenomegalia, pancitopenia, febre, caquexia e imunocomprometimento (6-8). A taxa de mortalidade entre os doentes de LV na maioria das áreas endêmicas é de 5-10% mesmo com tratamento específico. A maioria das infecções por L. infantum é resolvida espontaneamente. Estudos realizados no Brasil,na década de 80, demonstraram que cerca de uma em seis crianças infectadas por L. infantum desenvolve LV (9-11). No entanto, estudos mais recentes indicam que a proporção de pessoas infectadas que adoecem aumentou, potencialmente devido a fatores de risco associados ao desenvolvimento de LV, como nutrição e co-morbidades (12;13). Um marcador consistente de cura da infecção e proteção contra subseqüente re-infecção é o teste cutâneo de hipersensibilidade tardia (Delayed-type Hypersensibility DTH) ao antígeno de Leishmania, também conhecido como teste de Montenegro (14;15). Fatores que determinam se uma pessoa resolverá espontaneamente a infecção e desenvolverá a resposta DTH, ou se 12
, ou em uma doença")
17 prosseguirá para a doença sintomática e potencialmente fatal são entendidos de maneira incompleta (16;17). A LV é endêmica em 88 países e 90% dos casos concentram-se em Bangladesh, Brasil, Índia, Nepal e Sudão, com a 2 milhões de novos casos por ano (18;19). Essa doença tende a ser fatal se não tratada e mesmo com tratamento poderesultar em 5-10% de óbito (20;21). No Rio Grande do Norte ocorreu uma epidemia de LV no início da década de 1990 quando a taxa de mortalidade chegou a 9%; seguido de periurbanização, com áreas endêmicas novas localizadas nos arredores das maiores cidades do estado (3;22). O agente etiológico causador da leishmaniose visceral no Brasil é a Leishmania chagasi, enquanto que na a Índia e no Sudão o agente causador é a L. donovani (23;24). Na Europa, o agente causador da LV é a L. infantum, que é geneticamente idêntica à L. chagasi (25), portanto, doravante iremos utilizar L. infantum para nominar a espécie circulante no Brasil. Infecção por outras espécies de Leishmania, como L. amazonensis e a L. tropica, que frequentemente causa doenças cutâneas pode também resultar em LV (26-29). O genoma das espécies de Leishmania do Velho Mundo (Complexos L. donovani e L. major) compreende 36 cromossomos, enquanto que o daquelas oriundas do Novo Mundo (complexos L. mexicana e L. brasiliensis) possuem 34 e 35, respectivamente (30;31). O seqüenciamento do genoma da L. major foi realizado, demonstrando que os 32,8 Mpb dessa espécie codificam 8395 genes para proteínas e 711 genes de RNAs. Os genes se agrupam em clusters e a expressão gênica pode ser regulada em diversos níveis, desde o rearranjo de histonas até as modificações pós-traducionais (32). Subsequentemente, o 13

18 sequenciamento dos genomas de L. infantum e de L. braziliensis encontrou 8195 e 8314 genes nos genomas dessas espécies. Apesar de milhões de anos de evolução separando esses dois grupos (Leishmanias do velho e novo mundos), foi encontrada sintenia de 99% dos genes das três espécies. A transmissão dos parasitos se dá através de picadas de fêmeas hematófagas de insetos da família Psychodidae, subfamília Phlebotominae, no momento do repasto sanguíneo (33). Parece haver especificidade entre a espécie do vetor e a espécie de parasito que ele transmite (34). No Brasil, insetos do gênero Lutzomyia são os principais vetores de Leishmania e no Rio Grande do Norte, Lu. longipalpis foi encontrado em 93% dos municípios estudados (35), mostrando a grande extensão potencial de áreas endêmicas para leishmaniose visceral. Esses vetores são infectados quando picam mamíferos, principalmente cães, que funcionam como reservatório (36). Há evidências de que seres humanos podem também ser reservatórios (37). Os vertebrados apresentam a forma intracelular da Leishmania, formas amastigotas, que estão normalmente presentes em células de origem monocítica ou em neutrófilos. No trato digestivo do mosquito, as formas amastigotas são convertidas na forma flagelada, promastigota. Os promastigotas passam por processo de diferenciação, originando as formas metacíclicas, formas infectantes, que são inoculadas na derme dos vertebrados quando do repasto dos vetores. A infecção se dá quando formas metacíclicas atingem a corrente sangüínea e lá se diferenciam na forma amastigota. Após inoculação na derme, as leishmanias são internalizadas por neutrófilos e células de origem monocíticas. No interior dessas células as formas metacíclicas se transformam em amastigotas, que se multiplicam no interior dos fagossomos (38). A Leishmania 14
.")
19 é capaz de infectar macrófagos do fígado, baço, medula óssea e linfonodos (39;40). Antígenos presentes na saliva do flebotomíneo podem alterar a resposta imune do hospedeiro à Leishmania. Trabalhos mostraram que a saliva de Phlebotomus papatasi aumentava a infecção por L. major, impedindo a ativação de macrófagos via interferon gama, diminuindo a produção de óxido nítrico (41). Além disso, a picada de P. papatasi elicita uma resposta DTH (Delayed-Type Hypersensitivity ou hipersensibilidade tardia), o que aumenta o fluxo sangüíneo local, diminuindo o tempo para o repasto (42). No entanto, trabalhos mais recentes relatam que proteínas encontradas na saliva dos flebotomíneos são capazes de elicitar uma forte resposta protetora no hospedeiro contra a infecção por Leishmania, tanto em modelos experimentais (hamsters) quanto em reservatórios naturais (cães). Tais proteínas podem inclusive ser candidatas a uso em vacinas (43). O quadro de manifestação clínica resultante da infecção por Leishmania depende de interações entre o parasita e o hospedeiro, de sua capacidade de sobrevivência no organismo do hospedeiro, de sua virulência e de fatores intrínsecos do hospedeiro (isto é, seu grau de resistência à infecção pelo parasita) (44-47). Fatores nutricionais parecem estar também envolvidos na susceptibilidade à doença (48;49). O diagnóstico da leishmaniose visceral é realizado primeiramente baseado em dados clínicos, epidemiológicos e laboratoriais. Os pacientes evoluem com queixas de astenia, adinamia e freqüentemente apresentam coinfecções por vírus, bactérias e/ou fungos. Apresentam anemia variável, hepatoesplenomegalia, leucopenia e aumento da velocidade de sedimentação 15
, o que aumenta o fluxo sangüíneo local, diminuindo o tempo para o repasto (42).")
20 de hemácias. O padrão ouro para o diagnóstico da LV ainda permanece na demonstração parasitológica, observação direta do parasito em aspirados de medula ou de baço. Existem exames sorológicos, (Enzyme-Linked Immunosorbent Assay - ELISA) ou moleculares (Polymerase Chain Reaction PCR) (50-53), mas estes ainda não estão disponíveis para todas as localidades endêmicas. O teste de hipersensibilidade tardia (Delayed type hypersentivity response) só apresenta resultados positivos após, em média, 6 meses após a cura clínica da leishmaniose visceral.. No Rio Grande do Norte, nas últimas duas décadas, a Leishmaniose Visceral, que era de ocorrência tipicamente de áreas rurais, passou a ser registrada na periferia das áreas metropolitanas devido a migração de famílias residentes no campo para as cidades, processo similar que ocorreu em diversos estados do Brasil, como Piaui, Ceará, Paraíba, Minas Gerais, entre outros.. Epidemia de LV foi registrada em Natal nofinal da década de 80, (3;54), e mais recentemente, na cidade de Mossoró. As demais cidades de porte menor do estado também têm relatado casos da doença, configurando assim a LV como uma doença de ocorrência em áreas periurbanas Resposta imune à Leishmania Após uma continuada estimulação antigênica, células T CD4 + podem se diferenciar em diferentes tipos de células efetoras, que se caracterizam pelos tipos de citocinas que expressam. Células T helper (Th) 1 são células especializadas na defesa contra vírus e outros parasitos intracelulares e expressam interleucina 2 (IL-2), IL-12, Interferom Gama (IFN- ) e Fator de 16

21 Necrose Tumoral (TNF- ). As células que se diferenciam em Th2 estão envolvidas em respostas alérgicas e na defesa contra helmintos, e produzem IL-4, IL-5, e IL-13 (55-57). O perfil Th1 ou Th2 é por ativação de sinalização intracelular mediada por via IL-12/STAT4 ou IL4/STAT6, respectivamente (58;59). Em camundongos infectados por L. donovani, quando células T CD4 + expressam IL-4 há uma exarcebação da infecção e, na presença de Fator de crescimento transformante beta (TGF- ), parece ocorrer os sintomas resultantes da visceralização do parasita (60). A capacidade de auto-resolução da infecção por Leishmania em camundongos está ligada à presença de células Th1, à positividade na resposta DTH e à presença de citocinas ativadoras de macrófagos (61;62). Murray e colaboradores (1982) demonstraram em camundongos que macrófagos peritoneais não estimulados não conseguem eliminar amastigotas de L. donovani, mas quando estimulados por linfocinas, essas células conseguem destruir mais de 85% dos amastigotas (63;64). A ativação de macrófagos na infecção por Leishmania spp. é desencadeada pela ação de citocinas, principalmente (INF- ) (liberado por células T e NK), fator de necrose tumoral (TNF- ) e IL-2 (65-69). Murray e colaboradores (1990) demonstraram em camundongos que o INF- estimula a atividade leishmanicida de macrófagos, recruta células mononucleares para o sítio de infecção e é um importante co-fator para a resposta ao tratamento com antimônio (70). O gene do IFN- é ativado pelo fator de transcrição NFAT (Nuclear factor of activated T cells), que também ativa os genes das citocinas IL-2, IL-4, IL-5 e TNF- (71). Foi demonstrado que macrófagos ativados por IFN- e TNF- utilizam-se de espécies reativas de nitrogênio (NO - ) e oxigênio 17
22 (H 2 O 2 ) para a destruição de amastigotas de Leishmania spp. (72-75). Essas espécies reativas são lançadas no fagossomo onde estão as amastigotas. Também há evidências de que o NO - atua na sinalização molecular, ativando IL-12, que por sua vez tem papel determinante na expressão da resposta do tipo Th1 (76). Pacientes com leishmaniose visceral têm a resposta Th1 diminuída, com baixos níveis de IL-2, IL-12 e INF- (77-80). Os pacientes apresentam imunossupressão antígeno-específica, com resultados negativos ao teste de Montenegro (DTH-). Há reversão da imunossupressão após a cura clínica e diminuição dos níveis de anticorpos (81). Por outro lado, se células TCD4 + expressam TNF-, IL2 e IL12, ocorre uma resposta protetora, com controle da infecção. O TNF- atua ativando, juntamente com INF-, a produção de espécies reativas de nitrogênio e oxigênio (82;83). É sabido, no entanto, que uma quantidade de parasitos persiste no hospedeiro infectando células dendríticas e macrófagos mesmo após a cura clínica da doença (84-88). Essa persistência de parasitos pode levar a episódios de reinfecção, principalmente em pacientes HIV-positivos ou em tratamento contra rejeição de órgãos transplantados, caso haja perda da imunidade (89-91). Recentemente foi demonstrado, tanto em camundongos quanto em humanos, a existência de um subconjunto de célula TCD4+ (5-10%), que apresentam em sua superfície os marcadores CD25 e CTLA-4. Essas células foram denominadas de Treg e expressam altos níveis de IL10 e TGF- (92;93). As células Treg estão envolvidas no controle de células T CD4 + autorreativas, atuando no controle de processos autoimunes (94-97). Entretanto, podem suprimir processos benéficos, como a expressão da resposta Th1, concorrendo 18
23 para a manutenção de células parasitárias que mantém o sistema imunológico em estado de constante atividade denominado imunidade concomitante (98-100). Em modelo murino de infecção por L. major as células T CD4 + CD25 + diminuem significativamente, in vitro, a produção de IFN- por células T CD4 + efetoras (101). A transferência de linfócitos Treg de camundongos C57BL/6 (resistentes à infecção) curados de leishmaniose cutânea para camundongos da mesma linhagem cronicamente infectados por L. major causa um aumento no número de parasitos nesses camundongos, mostrando o papel de Treg na reativação da doença (102). Essa reativação não passa pela polarização da resposta celular para o pólo Th2, como seria razoável imaginar. Na verdade células Treg suprimem também a resposta Th2 (103). Seu mecanismo de ação é independente e está associado à produção de IL-10 e TGF- (104). Os resultados dos estudos acima revistos demonstram a complexidade da resposta imunológica à infecção por Leishmania spp. Intrincadas interrelações acontecem entre diferentes tipos celulares, com a produção de diversos tipos de citocinas que, por sua vez, influenciam a produção umas das outras, assim como a produção de espécies reativas de oxigênio e nitrogênio. Portanto, o grau de resistência ou susceptibilidade pessoas frente à infecção por Leishmania spp. deve ser determinado de maneira complexa, dependendo da atuação de vários dos genes que codificam proteínas que fazem parte da resposta imune, cada um com pequeno grau de contribuição para a resistência/susceptibilidade. Deve-se ainda levar em consideração as diferenças interpessoais e interpopulacionais nas seqüências dos genes envolvidos na resposta imunológica, já que esses genes estão submetidos aos 19
24 fatores evolutivos tais como seleção, mutação, deriva e recombinação genética. 2.3 Variação genética na resposta imune: As citocinas que participam da resposta imunológica, assim como seus receptores, são proteínas. Os genes que as codificam estão sujeitos a polimorfismos em suas seqüências que podem levar a variações no nível de expressão das citocinas ou seus receptores e, conseqüentemente, no tipo da resposta imune elicitada em resposta a um estímulo, no caso à infecção por Leishmania. Diversos estudos têm sido conduzidos no sentido de verificar a associação de polimorfismos em genes envolvidos na resposta imunológica à susceptibilidade de indivíduos a uma série de doenças como doenças auto imunes, por exemplo, lupus eritematoso sistêmico, e alérgicas, como asma e também à infecção por parasitos intracelulares ( ) e à progressão de doenças (112). As variações genéticas (polimorfismos) estão sendo anotadas em bancos de dados internacionais tais como NCBI ( GeneDB ( e Ensembl ( e por grupos que pesquisam especificamente variações em genes envolvidos na resposta imunológica (113;114). Polimorfismos de uma base (Single Nucleotide Polymorphisms) - SNPs - são polimorfismos bastante utilizados atualmente em estudos de associação, na busca do entendimento da atuação de fatores genéticos em doenças complexas (115;116). Um SNP por si só, não é informativo, mas quando 20
25 analisados em conjuntos, esses polimorfismos formam haplótipos, que por sua vez podem ser avaliados com relação à associação com quadros clínicos. 2.4 Etnia e susceptibilidade a leishmaniose visceral Brasil e Sudão O Brasil foi primeiro habitado por tribos de Ameríndios desde cerca de anos atrás, (117;118). Essas tribos se originaram a partir do fluxo de indivíduos asiáticos que atravessaram o estreito de Bering. Essa travessia provavelmente constituiu um gargalo genético, devido às condições climáticas restritivas. Ao longo do tempo o fluxo migratório continuou rumo ao sul, de modo que, os Ameríndios contavam com uma população estimada de 2.4 milhões de indivíduos no século XVI (119;120).No ano de 1500 os portugueses chegaram à região Nordeste do Brasil, seguidos pelos franceses e outras nacionalidades. Estes passaram a viver na terra nova e a socializar amigavelmente com os índios; Os portugueses começaram a exploração do Brasil pelo ano de 1530, com o envio de pessoas, a maioria do sexo masculino. Entre os séculos XVII a XIX, o Brasil recebeu levas de escravos africanos comprados pelos europeus de escravizadores pertencentes aos grandes reinos africanos (121). Os escravos eram provenientes de vários países, principalmente Guiné, Costa do Marfim, Congo, Angola e Benin (122;123). Cerca de quatro milhões de escravos foram trazidos e os principais portos que os receberam foram Salvador, Rio de Janeiro, São Luís e Belém. Os descendentes dos africanos contribuíram para o pool gênico que formou a população brasileira (122). 21
26 Devido ao modo como a colonização do Brasil foi conduzida, não se pode esperar que tenha havido panmixia entre os diversos grupos étnicos envolvidos. Homens europeus, principalmente portugueses, devem ter contribuído mais que homens africanos e ameríndios no processo reprodutivo. De fato, vários trabalhos mostram que este é o caso ( ). Em 1999 Batista dos Santos e colaboradores mostraram, em uma população de Belém, estado do Pará, que marcadores de DNA mitocondrial eram predominantemente ameríndios, enquanto que para o marcador da porção nãorecombinante do Y (DYS199) o alelo específico para populações ameríndias estava presente em apenas 3,8% os indivíduos analisados. Por outro lado, o alelo europeu estava presente em 96,2% dos indivíduos (128). Foi demonstrado que uma população afro-descendente semi-isolada, também na região amazônica é heterogênea com relação às contribuições materna e paterna: cromossomos Y têm origem predominantemente européia e DNA mitocondrial tem origem africana e ameríndia (129). Nós esperamos que este padrão se repita no Nordeste do Brasil. Entretanto, escravos que vieram para o Rio Grande do Norte (Leste do Nordeste) podem ser diferentes daqueles que foram levados para o Pará e Maranhão (Norte do Nordeste) e para a Bahia (Sul do Nordeste). Analisando a população brasileira como um todo, Carvalho-Silva e colaboradores (2001) demonstraram que a participação paterna de ameríndios para a população atual é quase nula e que há apenas uma participação muito pequena de africanos no Norte do país (130). Em todo o país predomina a descendência européia e em menor grau asiática e africana. A região Sul é mais heterogênea, devido as diversas nacionalidades que migraram para os diversos estados no século XIX e início do século XX (131). 22
27 O fluxo gênico é um dos principais fatores de variação em freqüências alélicas e genotípicas (132). Pessoas oriundas de populações indígenas foram miscigenadas com pessoas de diferente background genético, após a colonização do Brasil. Esses três principais grupos étnicos foram os formadores da população nordestina inicial e possuem diferenças genéticas (p. ex. marcadores de cromossomo Y) e fenotípicas (características físicas) que podem ser utilizados para questões de agrupamento étnico. No nosso trabalho usamos marcadores genéticos para estimar o percentual de formadores étnicos da população atual. Estudos genéticos em populações como a brasileira são dificultados pelo alto grau de miscigenação existente. Por isso, é importante estratificar a população nos seus grupos étnicos originais. Entretanto, essa tarefa não é simples porque, além do próprio fluxo gênico entre diferentes etnias ao longo do tempo, também pode ocorrer recombinação genética nos diversos loci entre gerações. A determinação de etnias constituintes de populações pode ser realizada de diversas maneiras. No Brasil há a prática de usar caracteres como cor de pele, cor e textura dos cabelos e forma do nariz e lábios para diferenciar pessoas por raça. Porém, já foi demonstrado que há pouca correlação entre esses traços fenotípicos e origem étnica (133). A atribuição de etnia deveria ser feita geneticamente, de maneira a evitar o viés introduzido por representações sociais artificiais. Polimorfismos em genes envolvidos na resposta imunológica já são bem conhecidos e há trabalhos mostrando alelos associados à susceptibilidade a doenças infecciosas em diferentes populações ( ). Predominam os trabalhos com tuberculose, e há poucos estudos que avaliam associações 23
28 entre polimorfismos e susceptibilidade à leishmaniose visceral. Diversas doenças têm incidências variáveis entre diferentes grupos étnicos ( ). A susceptibilidade a doenças também, aparentemente, pode ser influenciada por diferenças étnicas. Ibrahim e colaboradores (1999) demonstraram diferenças de susceptibilidade à leishmaniose visceral entre membros de diferentes tribos sudanesas, com incidência de LV significativamente mais altas em algumas tribos (140). Análises de polimorfismos no gene SLC11A1, envolvido no transporte de íons divalentes no retículo endoplasmático de macrófagos (141), demonstraram associação entre alelos presentes na etnia Masalit e susceptibilidade à LV (142;143) indicando a etnia como fator influenciador na susceptibilidade. Recentemente Miller e colaboradores (144) demostraram, em um estudo de varredura de genoma, que linhagens de populações como determinadas por polimorfismos no cromossomo Y estavam associadas a regiões cromossômicas no genoma autossômico em populações de duas vilas da etnia Masalit, susceptível à leishmaniose visceral. Isso se deu porque nesse caso em particular a quantidade de casamentos consangüíneos é grande. A genotipagem de polimorfismos no cromossomo Y permitiu a criação de três haplótipos, um dos quais marcava diferentes regiões cromossômicas nas duas populações, evidenciando um possível efeito fundador na criação das duas vilas (145). A escolha e utilização de marcadores genéticos da região nãorecombinante do cromossomo Y facilita a análise do componente genético da susceptibilidade, já que elimina o efeito da recombinação genética sobre a análise dos dados. Devido à miscigenação, o desenho de estudo de escolha é o estudo de associação baseado em famílias, que elimina o efeito de 24
29 estratificações na população. A determinação da influência ou não da etnia na susceptibilidade à leishmaniose visceral auxiliará no entendimento da dinâmica da doença no Estado e contribuirá para o conhecimento do funcionamento do sistema imunológico frente a infecções por parasitos intracelulares. 25
30 3. Material e métodos 3.1. Varredura de genoma Área de estudo Os participantes do estudo foram recrutadas entre pessoas residentes no na região leste do estado do Rio Grande do Norte, na área perimetropolitana ao redor da capital, locais endêmicos para LV. A cidade de Natal e as áreas vizinhas têm sido endêmicas para LV desde meados da década de 1980 (146) sendo responsável por 70% de todos os casos relatados no estado entre 1990 e A LV se concentra em pequenos focos geográficos e as taxas de infecção variam com o tempo em cada vizinhança. Foram identificadas vizinhanças com transmissão de L. infantum em curso através de casos de LV admitidos em hospitais públicos de Natal Exames físicos e estudos laboratoriais Após obtenção do consentimento livre e esclarecido, os participantes com LV e seus familiares foram entrevistados sobre seus históricos médicos, relações familiares e exposições ambientais. Amostras de sangue foram coletadas para determinação de contagem de células sanguíneas e o diferencial celular, hematócrito, sorologia para Leishmania e outros parâmetros quando indicado por razões médicas. O DNA foi extraído de leucócitos 26
31 conforme descrito por Karplus et al., 2002 (147). Anticorpos contra L. infantum foram detectados usando ensaio sorológico por ELISA (Enzyme-linked immunosorbent assay) com a cepa brasileira de L. infantum MHOM/BR/00/1669 como fonte de antígenos (148;149). A resposta DTH foi avaliada usando 25ug de antígenos de L. infantum gentilmente cedidas pelo Dr. Steve Reed (Instituto de Pesquisa de Doenças Infecciosas, Seattle, EUA). Após 48 a 72 horas da aplicação do teste, a área de enduração foi medida. Enduração igual ou maior que 5 mm é considerada uma resposta positiva indicando exposição anterior a Leishmania (150) Famílias e ambiente As famílias recrutadadas no estudo incluíram aquelas com casos de LV ou vizinhos que estivessem vivendo na área endêmica por no mínimo 2 anos. Desta maneira, alguns pessoas estavam infectados com Leishmania (determinado pelo teste DTH), mas não possuíam parentes com histórico de LV. Além disso, de acordo com registros do banco de sangue, a taxa de infecção por Trypanossoma cruzi (que pode ser um agente confundidor para o teste de DTH) é baixa na região de estudo. O estudo de marcadores étnicos utilizou os mesmos participantes da pesquisas dos projetos avaliados pelo CEP-UFRN e CONEP. Especificamente: CEP-UFRN 19-01, CONEP 4572; CEP-UFRN 20-01, CONEP 4575; CEP- UFRN 21-01; CONEP 4581; CEP-UFRN 94-04, CONEP
32 Definições dos fenótipos Os fenótipos considerados no presente estudo foram aqueles definidos em estudo realizado anteriormente (Jeronimo et al, 2004), sendo estes baseados na presença de história de LV ou após a interpretação dos resultados que incluíram a avaliação clínica, presença ou não de anticorpo anti-leishmania e/ou o tipo de resposta frente a intradermo reação (teste de Montenegro). Portanto, foram considerados os seguintes fenótipos. 1. Fenótipo Leishmaniose visceral (LV): o diagnóstico de LV foi baseado na história clínica característica de LV (presença de sintomas e epidemiologia), com validação laboratorial como esfregaço de medula óssea positivo para o parasito ou sorologia anti-leishmania positiva, e resposta ao tratamento (151). Pessoas com LV incluíram aqueles com doença sintomática na época do arrolamento ou histórico de doença validado pela revisão de prontuários médicos. 2. Fenótipo DTH+ (Montenegro positivo): o grupo DTH+ incluiu apenas pessoas sem histórico de LV sintomáticae uma reação de Montenegro positiva quando o tamanho da enduração é 5mmO tamanho da enduração, em mm, foi usado como caráter quantitativo.quando repetidos, os testes de Montenegro foram aplicados com intervalos de um ano para evitar boosting (152).. 3. Fenótipo com Sorologia positiva (DTH- Ac+): esse grupo incluiu 56 dos 107 indivíduos com sorologia positiva para Leishmania e resposta negativa do teste de DTH. Para as análises genéticas os indivíduos Ac+ foram considerados como de fenótipo desconhecido já que a sorologia positiva é indicativa de infecção aguda (153), a qual pode progredir para os fenótipos LV, DTH+ ou DTH- ( ). 28
33 4. Fenótipo Teste cutâneo e sorologia negativos (DTH-): essa categoria incluiu pessoas com sorologia anti-leishmania negativa e enduração < 5mm no teste de Montenegro que vivesse na mesma residência que sujeitos com VL ou em vizinhanças com taxa de infecção de pelo menos 40%). DTH- foram considerados possivelmente expostos a Leishmania embora à primeira análise não apresentassem evidência de resposta imunológica ao parasito. Foram excluídas dos estudos genéticos 3 pessoas que apresentaram infecção por HIV e quatro de seus parentes, além de pessoas que após a análise genética mostrou discrepâncias no grau de parentesco. Após essas exclusões 1254 indivíduos foram incluídos nas análises genéticas Genotipagem dos marcadores microssatélite: A genotipagem foi realizada no Centro para Pesquisa de Doenças Herdadas (Center for Inherited Disease Research CIDR Universidade Johns Hopkins, Baltimore, Maryland, EUA) com permissão do Conselho Nacional de Pesquisa (RMX 18/04). Marcadores STR (Short Tandem Repeats) distribuídos ao longo do complemento autossômico do genoma humano foram genotipados em todos os indivíduos. Esses marcadores (402 no total) são uma modificação do conjunto de varredura Marshfields Genetics versão 8, com distancia genética média de 9cM, sendo que não houve espaços maiores que 20cM. A taxa geral de perda de informação da genotipagem foi de 5,54% Controle de qualidade dos dados: Os dados foram liberados pelo CIDR após execução do programa GAS (Genetic Analysis System Sistema de Análise Genética) que identifica 29
34 inconsistências mendelianas e/ou problemas sistemáticos de manipulação no laboratório ou problemas de chamada dos alelos. Erros de relacionamento também foram checados pelo programa RELCHECK (157), o qual examina pedigrees grandes procurando, ao longo do genoma, padrões de compartilhamento de alelos entre classes de parentes. Também foram identificados quaisquer erros mendelianos através da utilização da opção error do programa MERLIN (V1.0 alpha) (158). Todos os erros foram corrigidos antes das análises. As freqüências dos alelos dos marcadores foram calculadas apenas para os fundadores e todos os marcadores estavam em equilíbrio de Hardy-Weinberg (EHW) Análises estatísticas da varredura de genoma: - Herdabilidade e análise de ligação quantitativa: o tamanho da enduração no teste cutâneo foi transformado em logarítmo para normalizar os dados para análise do caráter DTH como quantitativo. O fenótipo DTH logaritmizado tem distribuição normal, determinado pelo gráfico quantil-quantil. Todos os indivíduos classificados como LV foram codificados como de fenótipo desconhecido para DTH já que o tamanho da enduração varia em função do tempo após a doença. Correlações familiares foram estimadas utilizando o programa FCOR do SAGE (Statistical Analysis for Genetic Epidemiology v 4.8). A herdabilidade (h 2 ), que é a proporção da variação que é atribuída a fatores genéticos aditivos, foi estimada a partir de correlações entre irmãos (r) usando a equação h 2 =2r. Foram realizadas duas análises de ligação com o fenótipo contínuo DTH usando o programa MERLIN (V1.0 alpha) (158). Primeiro, foi feita análise de 30
35 regressão quantitativa não-paramétrica para testar o compartilhamento de alelos entre indivíduos com respostas DTH semelhantes. Aqui a média da amostra foi usada para estimar a média fenotípica, e o grau de compartilhamento de alelos por identidade por descendência (IBD Identical by Descent) foi estimado por métodos de análise multiponto usando famílias completas no MERLIN (158). Esse programa faz análises de ligação de componentes de variância sob a prerrogativa de que não há dominância e calcula a herdabilidade específica de cada marcador para cada caracter (159). - Análise de ligação qualitativa: para o fenótipo dicotômico LV foi realizada análise de ligação não-paramétrica no MERLI N. Foi testado se havia excesso de compartilhamento de alelos IBD entre parentes afetados usando a estatística S-all originalmente descrita por Whittermore e Halpern (2006) (160). LOD escores para o compartilhamento de alelos foram também calculados usando o modelo linear de Kong e Cox (1997) (161). Para todas as análises de ligação, calculamos valores p nominais através de todo o genoma. Para os marcadores STR dando evidência sugestiva de ligação (valor p < 0,001 para a regressão dos caracteres quantitativos e análise de ligação de componentes de variância) foram calculados os valores p empíricos para cada cromossomo usando simulação de perda de genes no programa MERLIN. Sob essa condição de simulação, genótipos aleatórios foram gerados para cada marcador condicionados às freqüências alélicas estimadas, padrões observados de dados perdidos e mapas genéticos especificados sob a hipótese nula de não ligação ou sob a hipótese alternativa de associação ao fenótipo (162). Para cada cromossomo com uma região de interesse, replicatas foram simuladas. 31
36 Considerações éticas Consentimento por escrito foi obtido de adultos e pais ou guardiões legais de menores de 18 anos. Jovens de 12 a 17 anos também deviam consentir por escrito. Protocolos de pesquisa foram aprovados pelo Comitê de Ética em Pesquisa da Universidade Federal do Rio Grande do Norte (nº e 21-01) e Comissão Nacional de Ética em Pesquisa (nº 4581 e 4575), Universidade de Iowa, Universidade Johns Hopkins, Universidade da Virgínia e Instituto Nacional de Pesquisa do Genoma Humano, Institutos Nacionais de Saúde, Estados Unidos. Sujeitos com problemas de saúde descobertos durante o estudo foram tratados pela equipe de campo ou remetidos à fonte médica apropriada em Natal Estrutura étnica de populações do Norte e Nordeste do Brasil e reanálise dos marcadores da varredura de genoma da população de Natal RN Áreas de estudo Foram também estudadas pessoas residentes no Maranhão, Piaui e Pará, que foram recrutadas em virtude de em suas famílias terem sido registrados casos de LV, hanseníase ou tuberculose ou leishmaniose, enquanto na Bahia foram recrutadas pessoas com leishmaniose tegumentar (cutânea e mucosa) e no Rio Grande do Norte pessoas com LV.Todas essas doenças são causadas por microorganismos intracelulares. A Figura 1 mostra os estados de onde foram recrutadas as pessoas consideradas neste estudo. 32
37 Os estados do Piauí e Maranhão serão referidos como Norte_NE (NN); sob a denominação de Pará (PA) há pessoas de várias localidades (Belém, Marajó, Santarém, Monte Alegre e Igarapé-Mirim); as pessoas do Rio Grande do Norte (RN) foram recrutadas no litoral leste do estado, conforme caracterizado acima,e aquelas da Bahia (BA) são provenientes da do distrito de Corte de Pedra, município de Presidente Tancredo Neves. Figura 1: Mapa do Brasil indicando os estados de onde foram recrutadas as pessoas participantes no presente estudo. 33
38 3.2.2 Participantes Foram genotipados marcadores de etnia do cromossomo Y de 1019 pessoas do sexo masculino oriundas de 543 famílias nucleares, provenientes de diversas áreas endêmicas, de acordo com a tabela Rio Grande do Norte Os participantes do estudo residem na área perimetropolitana de Natal e outro municípios do litoral leste, sendo a maioria oriunda dos municípios denatal, Parnamirim, Extremoz e São Gonçalo. Os participantes tiveram sangue periférico coletado em tubos contendo EDTA para extração de DNA. Foram também entrevistados para a determinação dos heredogramas de cada família. A descrição completa do estudo foi anteriormente publicada (Jeronimo et al, 2004). Os dados dos participantes (identificação da família, identificação pessoal, identificação do pai, da mãe, sexo e fenótipo) foram armazenados em no programa Microsoft Excel 2003 (Microsoft Corporation) e heredogramas gerados no programa Progeny 7 (Progeny Software) O estudo de Belém Os indivíduos correspondentes às áreas denominadas Norte do Nordeste e Pará foram recrutadosatravés do Estudo de Famílias de Belém, conforme descrito em Blackwell et al., 1997 (163). Famílias com casos de hanseníase e tuberculose foram arroladas na cidade de Belém e aquelas com casos de leishmaniose visceral foram arroladas em Santarém, Teresina e São Luís. As famílias de Belém e Santarém pertencem à área denominada Pará e as de Teresina e São Luís pertencem à área Norte do Nordeste. 34
39 Bahia Sessenta e oito famílias foram arroladas para este estudo, metade das quais tinha pelo menos um indivíduo doente de leishmaniose cutânea ou mucocultânea. Todas vivem na área endêmica do município de Corte de Pedra, BA a 280 km de Salvador. O estudo tinha como objetivo verificar o agrupamento familiar de casos de leishmaniose muco-cultânea em uma região livre de L. infantum, mas endêmica para L. brasiliensis (164). Tabela 1: Origem das famílias participantes do estudo com a respectiva indicação dos tipos de doenças relacionadas ao recrutamento inicial Origem Participantes (n) Famílias nucleares (n) Doenças PA Hanseníase e tuberculose NN Leishmaniose visceral RN Leishmaniose visceral BA Leishmaniose mucocutânea TOTAL Genotipagem de marcadores do cromossomo Y Trinta e nove polimorfismos de um nucleotídeo (SNPs) da região nãorecombinante do cromossomo Y foram agrupados em cinco painéis e genotipados (Quadro 1). As PCRs foram feitas por Touchdown usando primers desenhados como publicados na literatura (165;166). A genotipagem foi realizada pelo método SNaPshot (Applied Biosystems), que consiste na amplificação por reação em cadeia da polimerase (PCR) de trechos do genoma que contém os marcadores. Os produtos de PCR são usados como molde 35
40 numa reação de sequenciamento quando há adição de um único didesoxirribonucleotídeo marcado com fluoróforo na posição do sítio polimórfico. Nessa última reação é usado um outro oligonucleotídeo característico para cada marcador. Esse oligonucleotídeo usado na reação SNaPshot anela-se ao produto de PCR,antes do sítio polimórfico, de modo que quando a reação ocorrer, a DNA polimerase incorporará 1 nucleotídeo exatamente correspondente ao sítio polimórfico (nucleotídeo na fita complementar). Como a reação de SNaPshot utiliza didesoxirribonucleotídeos, após a colocação do primeiro nucleotídeo a reação de síntese pára e o produto de reação é o oligonucleotídeo de SNaPshot mais um nucleotídeo marcado. A técnica permite a genotipagem de até 10 marcadores simultaneamente e os fragmentos são separados por eletroforese em capilar (minisseqüenciamento). Quadro 1. Painéis de marcadores do cromossomo Y utilizados (167) Painel Marcadores 1 M168, M45, M170, M89, M35, M3, M9, M173 2 M181, M220, M148, M174, M40, M201, M33 3 M112, M150, M02, M81, M123, M78, M41, M216 4 M172, M52, M72, M223, M26, M21, M62, M12 5 M269, M70, M17,M199, M19, M194, M214, M122 A Figura 2 mostra os haplogrupos e haplótipos determinados pela presença do alelo variante de cada marcador escolhido, de acordo com a 36
41 nomenclatura do Y Chromosome Consortium (YCC) (168). Não conseguimos determinar os haplótipos mais específicos para algumas das famílias estudadas, mas este fato não contribui negativamente porque desejamos separar as famílias em grupos étnicos grandes (origem continental - África, América e Europa). Figura 2: Relações entre os marcadores étnicos de cromossomo Y utilizados no presente estudo. São mostrados os marcadores que definem os ramos e os haplogrupos e haplótipos são mostrados no lado direito da figura. Círculos e barras são coloridos da seguinte maneira: pretos ramos africanos; laranjas ramos europeus; vermelhos ramo ameríndio. 37
42 Análise de etnia Dados obtidos com a técnica de SNaPshot foram analisados com uso do programa GeneMapper v 3.5 (Applied Biosystems). Após inspeção manual dos genótipos para evitar erros introduzidos pelo software, os dados foram exportados para uma planilha do Excel (Microsoft Corporation) e os haplogrupos e haplótipos assinalados manualmente de acordo com a nomenclatura publicada pelo (YCC) (169;170). Haplótipos foram carregados no programa Arlequim v3.11 (171) e nele foi realizada contagem de freqüência de haplogrupos, análise de variância molecular - AMOVA - (172;173) e índice de diversidade genética (Fst) (174) com permutações Comparação das frequências de grupos étnicos entre diferentes fenótipos associados à infecção por L. infantum. Os fenótipos utilizados foram aqueles definidos na análise de varredura de genoma como VL, DTH+ e DTH-AC- (abreviadamente DTH-). Analisamos um indivíduo por família nuclear, de maneira que cada um não era relacionado com os demais, em um total de 124 indivíduos. Verificamos as diferenças de frequência dos grupos étnicos africano e europeu, entre os fenótipos agrupados dois a dois (LV x DTH+ e LV x DTH-) através do teste exato de Fisher com iterações. Um p-valor menor que 0,05 foi considerado significativo. O grupo étnico ameríndio não foi utilizado por causa do diminuto número de indivíduos pertencentes a esse grupo. 38
43 Análise de associação dos dados da varredura de genoma em função dos estratos étnicos A varredura de genoma realizada na população de Natal RN foi um estudo de ligação genética (linkage) (175). Cento e trinta e um pessoas não relacionadas que foram genotipados naquele estudo e também foram genotipados para a investigação de etnia foram incluídos nessa nova análise, que é um estudo de associação. Todos tinham seus fenótipos determinados, sendo eles: LV (presença ou história de infecção sintomática por L. infantum); DTH+ (reação positiva ao teste de Montenegro, sem sintomas de Leishmaniose) e DTH- AC- (ausência de resposta imunológica à Leishmania). Dos 402 marcadores utilizados na varredura, escolhemos 19 presentes nas regiões indicadas como tendo ligação com a LV ou infecção assintomática: os marcadores dos picos de LOD score (D9S1118, D15S657 e D19S246) e marcadores upstream e downstream destes (Quadro 2). Também escolhemos 3 marcadores do cromossomo 22 que foi identificado como tendo associação com LV no extrato de origem europeia da população do RN (176). Foi realizado o teste de equilíbrio de Hardy-Weinberg usando o software PEDSTATS (177) dos 22 marcadores para a verificação da ação de fatores evolutivos na distribuição das freqüências alélicas. Foi verificado se havia diferença nas frequências alélicas dos marcadores comparando doença versus infecção assintomática (LV x DTH+) e doença versus não-infecção (LV x DTH-AC-) na população como um todo e nos estratos africano e europeu (separados em grupos em função dos haplótipos de cromossomo Y). O estrato ameríndio não foi analisado devido ao reduzido número de pessoas. Para avaliar a associação foi usado o teste do 39
44 qui-quadrado com correção de Fisher, com iterações (SPSS v17.0). Um p-valor menor que 0,05 foi considerado significativo. Alelos com freqüência menor que 5% foram analisados em conjunto, sendo denominados alelo 1. Quadro 2: marcadores genéticos utilizados na análise de associação com desfechos da infecção por L. infantum em função da origem patrilinear dos sujeitos de pesquisa. Cromossomos D9S1978 D15S655 D19S433 D22S692 D9S925 D15S652 D19S245 D22S445 D9S1121 D15S657* D19S178 D22S1169** D9S1118* D15S966 D19S246* D9S301 D15S642 D19S589 D9S1122 D9S922 D19S254 * Marcadores dos picos de LOD score dos cromossomos; ** Marcador do pico de associação com LV em populações européias 40
45 4. Resultados e discussão 4.1 Varredura de genoma considerando a doença leishmaniose visceral A amostra deste estudo incluiu 1254 pessoas oriundas de 191 famílias. As características da população foram semelhantes àquelas previamente descritas e algumas foram incluídas em um trabalho anterior (178). A população do estudo incluiu membros de 130 famílias com pelo menos um caso de LV e 61 famílias sem casos de LV. Entre as pessoas com LV, 78 eram sintomáticas no momento da participação no estudo, enquanto os demais casos de LB tinham documentação doença.. Pessoas com LV tinham 42,5% de seus parentes infectados com Leishmania, enquanto que nas famílias sem LV 37,2% estavam infectadas com Leishmania de acordo com o teste de Montenegro (Tabela 2). Os dados demográficos e as médias de enduração do teste de Montenegro estão na tabela 3. Tabela 2: Número de famílias e participantes do estudo com os seus respectivos fenótipos resultantes da exposição à Leishmania infantum. As famílias foram recrutadas devido a história de LV, certificada por registros hospitalares ou por residirem próxima a um caso de LV.. Os tipos de fenótipos são explicados no texto. Famílias Total de Fenótipos Participantes LV n (%) DTH+ n (%) AC+ n (%) DTH- n (%) LV (n=130) 162 (18,2%) 312 (35%) 67 (7,5%) 351 (39,3%) 892 Controle (n=61) 0 (0%) 95 (26,2%) 40 (11%) 227 (62,7%) 362 Total (n=191) 162 (13,4%) 307 (32,5%) 107 (8,5%) 578 (46,1%) 1254 LV (leishmaniose visceral); DTH (delayed type hypersensitivity test); AC (anticorpo anti-leishmania) 41
46 Tabela 3. Características demográficas das 1254 pessoas oriudas do estudo de linkage considerando exposição à Leishmania infantum. Idade (ano) Outras características >15 Total Sexo N(%) Idade quando LV N(%) Idade quando arrolado N (%) Homem 109 (18.9%) 82 (14.2%) 90 (15.6%) 295 (51.2%) Mulher 117 (17.2%) 69 (10.2%) 106 (15.6%) 386 (56.9%) Homem 59 (57.3%) 9 (8.7%) 7 (6.8%) 28 (27.2%) Mulher 49 (83%) 6 (10.2%) 2 (3.4%) 2 (3.4%) DTH+ 14 (3.4%) 33 (8.1%) 59 (14.5%) 301 (73.9%) VL 88 (54.3) 28 (17.3%) 15 (9.25%) 31 (19.1%) AB+ 12 (11.2%) 13 (12.1%) 21 (19.6%) 61 (57%) DTH- 112 (19.4%) 77 (13.3%) 101(17.5%) 288 (49.8%) 576 (45.9%) 678 (54.1%) 103 (63.6%) 59 (36.4%) 407 (32.4%) 162 (12.9%) 107 (8.5%) 578 (46.1%) média da enduração do DTH (6.6) 13.1 (6.5) 14.5 (7.5) 13.9 (7.8) 13.8 (7.7) Montenegro (SE) 42
47 Foi realizada uma varredura ao longo do genoma humano em busca de ligação de marcadores genéticos e LV como caracter qualitativo e DTH como caracter quantitativo. A linha sólida na figura 3 representa evidência de ligação genética de marcadores próximos a genes que controlam a forma sintomática da LV para todos os cromossomos autossômicos. O conjunto de famílias estudadas incluiu 21 pares de irmãos afetados concordantes para LV e 161 pares discordantes para LV. Além disso, houve pares de parentes de segundo grau concordantes incluindo 2 pares avunculares e 5 pares de primos. Houve 58 pares de primos não discordantes e 93 pares avunculares discordantes para o fenótipo LV. A despeito do número bastante limitado de pares de parentes afetados, foi identificada uma região de interesse no cromossomo 9q perto do marcador D9S1118 (p = 0,003, LOD = 1,60; valor p empírico = 0,0034). A figura 3 mostra a região de interesse no cromossomo 9. Para o caracter quantitativo resposta DTH, houve 440 pares de irmãos com uma correlação de 42%, 212 pares avôs-netos com uma correlação de 26,5% e 90 pares de primos com uma correlação de 13%. Isto é consistente com um controle genético para a resposta DTH, em que parentes de primeiro grau têm uma correlação mais forte que parentes de segundo e terceiro grau. A herdabilidade estimada da resposta DTH foi de 84%, sugerindo um componente genético substancial para variação da resposta DTH. Houve várias regiões de interesse para o caráter quantitativo DTH. As linhas tracejadas na figura 3 mostram os resultados de ligação para a resposta DTH medida pelo Log do tamanho da enduração. Foram identificados picos de Lod scores no cromossomo 15 (D15S657, p = 0,003; LOD = 2,5; p empírico = 0,008) e cromossomo 19 (D19S246, p = 0,001; LOD = 1,93; p empírico = 43
48 0,029) (Tabela 4). LOD scores dos loci de caracteres quantitativos (QTL) revelaram um padrão similar (Tabela 4). Esta análise também identificou regiões de interesse potencial nos cromossomos 2 e 13. Interessantemente, quando a análise foi restringida a indivíduos com pelo menos um parente com VL, os picos de ligação nos cromossomos 9 e 13 aumentaram, a despeito do menor número de famílias (Tabela 4). Isto pode refletir uma exposição mais uniforme dos indivíduos incluídos nesta última análise, na qual cada família tinha um membro com doença. Genes candidatos dentro dos picos de ligação estão 44
 para cada cromossomo baseado no mapa genético de Marshfield. Gráficos multiponto dos LOD scores de LV e resposta imune DTH.")
49 Figura 3. Sumário dos resultados da varredura de genoma considerando os fenótipos LV e infecção assintomática por Leishmania (tamanho da enduração a antígenos de Leishmania). O eixo X representa a distância em centimorgans (cm) para cada cromossomo baseado no mapa genético de Marshfield. Gráficos multiponto dos LOD scores de LV e resposta imune DTH. A linha sólida mostra a análise de ligação não-paramétrica do fenótipo LV. A linha tracejada mostra o resultado da análise de components de variância do tamanho da resposta de Montenegro (DTH), ajustada para idade e gênero. 45
50 Tabela 4: Principais Resultados da análise de ligação em todo o genoma da resposta de Montenegro (DTH), com as análises correspondentes para LV nas mesmas regiões. Leishmaniose Visceral Resposta de Montenegro Resposta de Montenegro apenas em famílias LV Cr a Marcador mais próximo cm Análise de ligação nãoparamétrica 34 famílias Análise de regressão 145 famílias Componentes de variância 179 famílias Análise de regressão 98 famílias Componentes de variância 119 famílias P LOD P LOD P LOD P LOD P LOD 2 D2S D13S D15S D19S (73) a Cr cromossomo. cm- CentiMorgan localização do marcador mais próximo. O número de famílias reflete aquelas que foram informativas para a análise de ligação. São indicados tanto os valores p quanto os LOD Scores. 46
51 Quadro 3. Exemplicação de genes presentes na área do genoma ligado aos fenótipos LV ou a resposta DTH. São mostrados genes selecionados dentro do intervalo de 1 LOD abaixo do pico de ligação a VL no cromossomo 9 e a DTH nos cromossomos 2, 13, 15 e 19. Dados são baseados no genoma humano v 36.1 (Homo sapiens Genome Build 36.1). Os genes estão listados em ordem, com a posição do marcador listada na segunda coluna da tabela. Cromossomo 9 (LV pico D9S1118; região 21,900K-41,900K) Símbolo Local Descrição CDKN2A 9p21 Cyclin-dependent kinase inhibitor 2A (melanoma, p16, inhibits CDK4) CDKN2B 9p21 Cyclin-dependent kinase inhibitor 2B (p15, inhibits CDK4) IFNK 9p21.2 interferon, kappa D9S1118 ACO1 9p22-p13 aconitase 1, soluble TRBV20OR9-2 through 9p21 T cell receptor beta chain variable region genes and A (9 genes) TRBV29OR9-2 IL11RA 9p13 interleukin 11 receptor, alpha CCL27, CCL19, 9p13 chemokine (C-C motif) ligands 27, 19 and 21 (3 genes) CCL21 CD72 9p13.3 CD72 antigen (B cell marker) Cromossomo 15 (DTH pico D15S657, região 79,600,000k-telomero) CIB1 15q25.3-q26 calcium and integrin binding 1 (calmyrin) MAN2A2 15q26.1 Mannosidase SLCO3A1 15q26 solute carrier organic anion transporter family member 3A1 LOC q26.1 Ferritin light chain 1 homolog D15S657 IGF1R 15q26.3 insulin-like growth factor 1 receptor Cromossomo 19 (DTH pico D19S246, região 43,650K-telomero) LGALS family 19q13.2-q13.1 Galectins 7, 4, 13, 14 IL28B-IL28A-IL29 19q13.13 interleukins 28B, 28A, 29 FCGBP 19q13.1 Fc receptor TGFB1 19q13.1 transforming growth factor beta 1 APOE 19q13.2 Apolipoprotein E PTGIR 19q13.3 Prostaglandin I2 (prostacyclin) receptor CD37 19p13-q13.4 CD37 IRF3 19q13.3-q13.4 interferon regulatory factor 3 D19S246 CD33 19q13.3 CD33 (gp67) CNOT3 19q13.4 CCR4 subunit 3 FCAR 19q13.2-q13.4 Fc alpha receptor IL11 19q13.3-q13.4 interleukin 11 O desfecho da infecção por Leishmania depende de uma interação complexa entre o parasito, o vetor flebotomíneo e o hospedeiro final, com características individuais de cada contribuindo com as manifestações da doença. Este estudo focalizou os fatores genéticos do hospedeiro humano controlando o desfecho da infecção com L. infantum, a causa de leishmaniose visceral no Nordeste do Brasil. A população de estudo foi extraída do local de 47
52 um recente ressurgimento de LV na zona periurbana de Natal, a maior cidade no estado de Rio Grande do Norte, Brasil. A população de estudo incluiu 36% pessoas com infecção assintomática, 11% com LV sintomática e cerca de 50% de pessoas sem evidência imunológica de infecção por L. infantum. Marcadores de infecção foram a sorologia, a qual é positiva durante a infecção aguda, e o teste de Montenegro, um marcador de infecção assintomática ou sintomática, que geralmente está presente quando há diminuição da resposta humoral (179;180). Existiam pessoas DTH- vivendo em residências com pessoas com história atual ou passada de LV por, indicando que essas podiam, potencialmente, terem sido expostas a Leishmania. É sabido que pessoas com teste de Montenegro positivo podem reverter para o fenótipo negativo (181). Conseqüentemente, o grupo DTH - considerado no presente estudo pode ser constituído por um grupo heterogêno de pessoas e poderiam incluir pessoas que foram anteriormente DTH+. Devido a esta incerteza, foram considerados apenas o fenótipo VL e o tamanho da reação DTH na análise de ligação. Este é o primeiro estudo de ligação com leishmaniose assintomática (DTH+) conhecido até hoje e sua análise revelou 4 regiões de interesse, nos cromossomos 2, 13, 15 e 19. O tamanho da reação DTH parece refletir a efetividade da resposta imune tipo 1 à L. infantum e a reação DTH poderia ser uma medida do desenvolvimento da imunidade protetora. Esses resultados provêem informação intrigante nos loci que podem controlar a resposta imune à LV. A sugestão de evidência mais forte para ligação da resposta DTH ocorreu nos cromossomos 15 e 19, com picos menores adicionais nos cromossomos 2 e 13. Nós hipotetizamos que genes diferentes influenciam a propensão a 48
53 desenvolver a infecção sintomática ou assintomática, e que vários genes podem controlar a resposta imune. As análises secundárias apenas com as famílias contendo membros com LV produziram LOD scores mais altos para dois loci, sugerindo que fatores de susceptibilidade influenciam a resposta de Montenegro em algumas famílias. Embora nenhuma das regiões encontradas foram significantes usando a definição de Lander e Krugylak, mas alguns loci apresentam evidência sugestiva de ligação (p = 0,00074) e essas regiões necessitam de estudos mais refinados (182). O resultado da análise dos genes candidatos presentes nas regiões de ligação mostra vários genes ligados à resposta imune como TGFα no cromossomo 2, carreadores aniônicos solúveis 2, 13 e 19, e SMAD 9 no cromossomo 13. O pico do marcador no cromossomo 19 está perto de genes codificando galectinas, receptor Fc, proteína de ligação 4 a TGF-β1, subunidades de IL-28, apolipoprotreína E e receptor de prostaglandina. Ainda mais, para o subconjunto de famílias múltiplas de LV, foi identificado um pequeno pico de ligação à LV no cromossomo 9, o qual está perto da subunidade α do receptor de IL11, um agrupamento de genes da região variável da cadeia β de receptor de célula T, e três ligantes de receptores de quimiocinas. Um estudo mais aprofundado, chamado mapeamento fino (finemapping), está em andamento para testar se quaisquer dos genes candidatos identificados, ou outros genes nas regiões de ligação encontradas, influenciam a resposta imune à Leishmania ou o desenvolvimento de LV. Há outras duas varreduras de genoma para LV relatadas até hoje. A primeira focalizou residentes de uma vila sudanesa expostos a L. donovani. Este estudo descreveu uma evidência significativa de ligação da doença 49
54 sintomática com a região do cromossomo 22q12 (LOD score 3,50) em uma varredura de genoma de famílias com múltiplos casos de LV em uma única vila no Sudão. Um lócus secundário (22q22-23) foi também sugestivo de ligação em famílias com LOD scores negativos para o locus 22q12 (183). Curiosamente, outros estudos de indivíduos com LV e leishmaniose cutânea pós- calazar (PKDL) em uma região geograficamente próxima à do estudo anteriormente relatado não encontrou evidência de associação com os mesmos marcadores, mas encontrou associação de LV com os genes IL-4 e SCL11A1, que codificam respectivamente IL-4 e NRAMP1, e associação da PKDL com o gene codificando a cadeia α do receptor de IFN-γ (IFNGR1) (184). Uma varredura do genoma com LV em pedigrees com múltiplos casos no Norte do Brasil identificou três regiões cromossômicas de interesse (6q27, 7q11.22, e 17q11.2-q21.3), com LOD scores máximos de 1,08, 1,34 e 1,14, respectivamente. O mapeamento fino na região de interesse do cromossomo 17 sugeriu associação alélica com dois genes de quimiocinas (185). Um subconjunto de nossas famílias com LV foi incluído naquele estudo. No presente estudo, no qual foi focalizado o fenótipo DTH, mas também analisado o fenótipo LV, não replicou os achados dos estudos prévios de ligação do fenótipo LV nas populações brasileira ou sudanesa. As diferenças com relação ao estudo sudanês podem ser devidas às diferentes espécies de Leishmania (L. donovani no Sudão vs L. infantum no Brasil) e diferenças genéticas entre as populações sudanesa e brasileira. O desfecho diferente do nosso estudo contra o outro estudo de varredura em famílias com múltiplos casos de LV no Brasil pode refletir diferenças genéticas entre os hospedeiros, ou mais provavelmente, a 50
55 heterogeneidade genética do caracter LV. De fato, brasileiros formam uma população geneticamente miscigenada e diferenças regionais já foram documentadas entre brasileiros de diferentes estados (186;187). Também é provável que o número relativamente pequeno de pedigrees informativos com múltiplos casos de LV neste estudo tenha falhado em detectar todos os genes controlando o risco de LV em populações endêmicas Estrutura étnica: Através da genotipagem de marcadores genéticos SNPs presentes na porção não-recombinante do cromossomo Y fomos capazes de determinar as origens étnicas, pela linhagem paterna, de famílias do Norte e Nordeste do Brasil. Os grupos étnicos foram agrupados no geral em Ameríndios, Africanos e Europeus. A Tabela 5 mostra as freqüências dos diferentes grupos nas populações estudadas e as proporções de cada grupo estão mostradas na Figura 4. Houve grande variabilidade nos haplogrupos encontrados nas três populações, com uma variedade de haplogrupos, desde o A até o T, agrupados em 22 haplótipos (188). Não foram observadas famílias pertencentes aos haplogrupos C e D, que marcam Ásia e Oceania (C) e Ásia central (D) (189;190). Como esperado, não há uma distribuição homogênea nas freqüências desses haplogrupos ou de haplótipos dentro dos haplogrupos (Tabela 5). 51
56 Figura 4: Mapa do Brasil indicando as freqüências de cada grupo étnico nos locais estudados. Preto Africano; Vermelho Ameríndio; Amarelo Europeu. Tabela 5: Freqüências relativas (%) dos haplogrupos de cromossomo Y encontradas nas áreas estudadas. BA RN Norte_NE PA A-B 1,47 2,27-0,55 E* 5,88 3, E1b1a - 5,00 16,90 12,02 E1b1b1 4,41 1,82 4,23 5,46 E1b1b1a 4,41 7,73 1,41 6,01 G* 2,94 2,27 5,63 4,92 H or J 8,82 6,36 1,41 7,65 I* 1,47 5,00 9,86 4,37 I2b 1,47 2,27 1,41 1,64 J2 2,94 6,82 5,63 3,28 J2b 4,41 1,36 1,41 1,09 P* 19,12 2, Q1a3a 5,88 4,55 8,45 6,56 R1b1b2 35,29 41,36 26,76 33,33 R ,63 8,74 Outros 1,47 7,27 11,27 4,37 Total 100,00 100,00 100,00 100,00 52
57 De maneira geral, as freqüências observadas estão em acordo com os dados encontrados na literatura, que mostram uma maioria de cromossomos Y europeus (70-80%), cerca de 20% de cromossomos africanos e o restante de ameríndios (191;192) (Figura 4). Esses valores refletem o processo histórico de colonização do Norte e Nordeste do Brasil, no qual inicialmente apenas homens europeus (principalmente portugueses) se estabeleceram no Brasil. Posteriormente outras nacionalidades européias migraram para o Brazil, além de outras nacionalidades africanas que foram escravizadas e trazidas para o país, passando por similar processo de miscigenação. Ao considerarmos somente os grupos que marcam populações européias, observamos que o haplogrupo R1b1b2, encontrado em alta freqüência em Portugal e típico de populações do oeste europeu (193), foi encontrado em freqüências acima de 35%, chegando a 55,48% no RN (Tabela 6). Interessantemente, haplogrupos presentes em populações judias e em outros povos de línguas semíticas foram encontrados no nosso estudo. São os haplogrupos G, J, e haplótipo R1a1 ( ). Cromossomos do haplogrupo G foram encontrados em uma freqüência de 3,77% na BA a 8,16% no NN. O haplogrupo J, incluindo aqui J2 e J2b, foi observado em 10,98% dos cromossomos europeus do RN e 10,2% dos do NN (Tabela 6). J2 M172G é encontrado em 24% dos judeus sefárdicos e M17 R1a1 é encontrado em até 52% dos judeus levitas Ashkenazi (197). O alelo mutante deste último foi observado em 1,22% e 2,04% no RN e NN, respectivamente, e ausente na BA. Se forem levados em consideração aqueles cromossomos que não puderam ter haplótipos definidos e estão no grupo H ou J, a contribuição de 53
58 cromossomos judeus pode ser ainda maior. Haplótipos do haplogrupo J são encontrados em alta freqüência em populações na Rússia, Lituânia, Ucrânia, Bielorrússia, e Hungria. Entretanto, sabemos que nenhum desses povos participou da colonização do Brasil. Estes resultados dão suporte à idéia de que judeus convertidos ao cristianismo (Novos cristãos) vieram ao Brasil no início de sua colonização, fugindo da perseguição pelos cristãos na Europa. Diversos trabalhos mostram a contribuição importante da imigração de judeus na formação da população da península ibérica e, em particular, da população portuguesa ( ). Também é interessante notar que as maiores freqüências desses haplótipos estão no RN e NN. Tabela 6: freqüências absolutas e relativas dos haplogrupos e haplótipos europeus encontrados nas famílias estudadas. n (%) BA RN NN PA G 2 (3,8) 5 (3,0) 4 (8,2) 9 (7,1) H ou J 6 (11,3) 14 (8,5) 2 (4,1) 14 (11,0) I 1 (1,9) 11 (6,7) 7 (14,3) 8 (6,3) I2a (2,0) - I2b 1 (1,9) 5 (3,1) 1 (2,0) 3 (2,4) J2 2 (3,8) 15 (9,1) 4 (8,2) 6 (4,7) J2b 3 (5,7) 3 (1,8) 1 (2,0) 2 (1,67) K-M - 6 (3,6) 3 (6,1) - K-R 1 (1,9) (5,5) P 13 (24,5) 6 (3,6) - - PQR (4,1) - R (8,26) 16 (12,6) R1a1-2 (1,2) 1 (2,0) 1 (0,8) R1b1b2 24 (45,2) 91 (55,5) 19 (38,8) 61 (48,0) T - 6 (3,5) - - Total 53 (100) 164 (100) 49 (100) 127 (100) Com relação às linhagens africanas, o haplótipo E1b1a, caracterizado por M02 G, foi encontrado em 5,0% das famílias do RN, 16,9% das famílias do NN, 21,0% nas do PA e não foi observado na amostragem da BA (Tabela 5). M02 é restrito à África sub-saariana, não sendo observado na Europa 54
59 (201;202). Em 4,41%, 7,73%, 1,41% e 6,01% das famílias estudadas (na BA, RN, NN e PA, respectivamente) foi detectado o alelo T do marcador M78, que marca o haplótipo E1b1b1a. Este haplótipo tem ampla distribuição, sendo encontrado na Europa, no Oriente Médio e no Norte e Leste da África (203). Devido ao fato de que os haplótipos E2, E3, E1b1b1, E1b1b1a e E1b1b1b são encontrados tanto na África quanto na Europa, os deixamos fora da tabela 6. A presença destes haplótipos em nossa população é muito provavelmente devida à miscigenação entre portugueses e mouros do norte da África, que invadiram a Península Ibérica e a dominaram por um período de 800 anos, além da entrada de escravos comprados na África no período entre os séculos XV e XVI. Portanto, para fins de análise de variação genética, consideramos todas as famílias com haplótipos contidos no haplogrupo E como sendo de origem africana. As freqüências de cromossomos ameríndios (M3 C - haplótipo Q1a3a) variaram entre 5,88% (BA) e 8,45% (NN) (Tabela 5). O marcador M3 é encontrado quase exclusivamente nas Américas (204) e está associado a populações autóctones deste continente. Utilizamos 3 marcadores (M19, M199 e M214) para refinar a análise destas famílias e tentar obter uma maior precisão quanto à etnia dos ancestrais indígenas. Entretanto, nenhuma família analisada apresentou alelos variantes para quaisquer destes marcadores. Os dados obtidos nesse estudo são, para o nosso conhecimento, os mais refinados já obtidos para a região Nordeste. Outros trabalhos publicados têm uma resolução limitada e mais superficial de populações nordestinas (205;206). 55
60 4.3. Variação genética entre as populações Foi realizada uma Análise de Variância Molecular (AMOVA) (207;208) com o software Arlequim v3.11 (209) para estudar as diferenças nas distribuições dos haplogrupos e haplótipos encontrados dentro e entre as populações estudadas. Como visto na Tabela 7, a fração da variância entre populações é bem pequena (1,77%), com quase toda a variação dentro das populações (98,23%). Isto é provavelmente devido ao fato de que a maioria dos cromossomos Y é de origem européia e, destes, a maioria pertence a um único haplótipo (R1b1b2), como discutido anteriormente. Tabela 7: Análise de variância molecular dentro e entre as populações estudadas como um todo, e com relação aos haplogrupos e haplótipos africanos. Variação* Dentro das populações Entre as populações Populações como um todo 98,2% a 1,8% a Y Africano 89,9% b 10,1% b Y Europeu 97,1% c 2,9% c * p = 0 em todos os casos c Fst global 0,02888 a Fst global 0,01766 b Fst global 0,10086 Sabendo que os africanos que chegaram ao Brasil vieram de vários locais distintos (122;123), decidimos então verificar a variância apenas de haplótipos africanos e, para testar a afirmação de que a pouca variação entre populações é devida à grande influência do haplótipo R1b1b2 (ibérico), analisamos também os cromossomos europeus. Quando comparamos a diversidade levando em conta apenas os haplótipos derivados da África, observamos um aumento na variação entre populações de cerca de 5 vezes (10,09%, p=0,0) (Tabela 7). A variância entre populações européias foi semelhante à variação global (2,89%, p=0,0), corroborando a idéia de que a variância genética encontrada é realmente devida aos indivíduos de etnias africanas trazidos ao Brasil e a herança européia é mais homogênea. 56
61 Considerando estudos realizado por Rosemberg et al., 2002 no qual foi realizada uma análise de variância molecular (AMOVA) baseada nas 52 populações do Painel de Linhagens Celulares de Diversidade do Genoma Humano (HGDP-CEPH) a variação genética intra-populacional é sempre muito maior que a inter-populacional, isto é, a variação genética observada é sempre maior dentro de uma dada população que entre populações diferentes. O teste de AMOVA feito nas 6 populações africanas do painel indicam uma variação inter-populacional de 3,1% e intra-populacional de 96,9%. A amostra do nosso estudo apresentou maior variação dentro (10,1%) e menor entre as populações (89,9%). Em seu trabalho Rosenberg e colaboradores utilizaram marcadores STR, que são autossômicos e passíveis de recombinação, enquando que nós usamos haplótipos formados por marcadores da região não-recombinante do cromossomo Y. Nossos dados indicam que a variação entre as populações africanas pode ser maior do que o estimado. Para verificar o grau de diferenciação entre as populações utilizadas no estudo, foram realizados testes dos valores de distância genética entre pares de populações (Fst). Os resultados indicam que há pouca diferenciação entre as populações estudadas como um todo, mas, novamente, levando em consideração apenas os cromossomos derivados da África, há diferenças significativas (chegando a uma ordem de grandeza): a população da BA é mais distante geneticamente da Pará que das populações do RN e Norte_NE, como se observa pelo maior valor de Fst e valor p significativo. A população NN está geograficamente localizada entre RN e PA e não difere significativamente de nenhuma destas (Tabela 8). A população do RN difere significativamente da do PA tanto do ponto de vista global como do componente africano. Isso é 57
62 explicado pelo fato de que os negros que vieram para o RN eram provenientes de Angola, Gana e Benin principalmente através do porto de Salvador. Aqueles que chegaram ao Norte do país vieram de Guiné, Guiné-Bissau, Cabo Verde e Gana através dos portos de Belém (PA) e São Luiz (MA) (122;123).Novamente, analisando apenas o componente europeu, as distâncias são reduzidas, mas também significativas. Nas duas análises precedentes o componente ameríndio foi excluído porque apenas um haplótipo marcador deste grupo étnico foi encontrado e, portanto, não é possível avaliar a variação entre os grupos. Obviamente as nações indígenas ancestrais dessas populações são distintas, mas nosso estudo não atingiu a resolução necessária para diferenciá-las. Tabela 8: Valores de distância genética (F st ) entre as populações estudadas. BA RN NN PA Populações BA como um todo RN c NN c b Estrato africano Estrato Europeu a p < 0,01. b p > 0,05 c 0,05 > p > 0,01 PA a a b BA RN b NN b b PA a c c BA RN a NN b b PA a b c Os resultados da análise de variância molecular (AMOVA) e das distâncias genéticas entre pares de populações indicam que a variabilidade genética entre as populações estudadas, de modo geral, é pequena, o que é explicado novamente pela grande proporção de famílias de origem européia. Ao mesmo tempo, mostram que o componente africano é variado. Valores de F st de cerca de 0.2 indicam uma grande diferença entre populações ou 58
63 subpopulações. Entre os pares de populações BA x PA (0,21), NN x PA (0,22) e entre RN x NN (0,17) no extrato africano essa diferença foi acentuada. Isso reflete o fato de que os escravos trazidos para o Norte vieram de Guiné, Guiné- Bissau, Cabo Verde e Gana pelos portos de Belém (PA) e São Luiz (MA) os que vieram para o Nordeste vieram de Angola, Gana e Benin via o porto de Salvador (BA). É sabido também que a Bahia exportava escravos para vários estados do Brasil a partir do início de 1800 (122;123) Comparação das frequências de grupos étnicos entre diferentes fenótipos associados à infecção por L. infantum. Os indivíduos estudados neste trabalho podem ser agrupados em função da sua origem étnica patrilinear. Foi então verificado se há diferença na distribuição de etnias entre indivíduos de diferentes fenótipos. Como há três tipos de fenótipos, as comparações foram feitas dois a dois (LV x DTH+ e LV x DTH-). Não foram encontradas diferenças significativas nas distribuições de etnias entre os diferentes fenótipos, sendo LV x DTH+ p-valor = 0,459 e LV x DTH- p-valor = 0, Análise de associação dos dados de varredura do genoma com fenótipos relacionados à infecção por L. infantum em função da origem étnica A partir das informações de origem patrilinear dos indivíduos do estudo de Natal RN, nós realizamos um teste de associação entre 21 dos marcadores utilizados na varredura de genoma conduzida com população da área endêmica para leishmaniose visceral desta mesma cidade, além de marcadores presentes no cromossomo 22 cujas regiões estavam associadas à 59
64 LV. O teste exato de Fisher foi utilizado para verificar se há excesso de algum grupo étnico nos indivíduos quando comparamos os fenótipos LV x DTH+ ou LV x DTH-, isto é, doença versus infecção assintomática e doença versus nãoinfecção. As pessoas foram estratificadas por origem étnica em dois grupos: africanos ou europeus. Todos os marcadores estavam em equilíbrio de Hardy- Weinberg, exceto o D15S816, que não foi considerado para análises posteriores. A tabela 9 mostra que há diferenças significativas entre as frequências alélicas do marcador D9S925 entre LV x DTH+ e LV x DTH- apenas nas pessoas de origem europeia, com p-valores de para LV x DTH+ e 0,033 para VL x DTH-AC-. O marcador D22S1169 mostra diferença nas distribuições das frequências alélicas com um valor p próximo da significância (p = 0,079) quando comparando indivíduos doentes x infectados assintomáticos na população europeia. A tabela 10 mostra as frequências alélicas do marcador D9S925. É importante salientar que este é um estudo de associação utilizando marcadores microssatélites, que são muito variáveis. O mais adequado neste caso seria a utilização de marcadores menos polimórficos, como os bialélicos. Ainda assim obtivemos um sinal de associação em um marcador no cromossomo 9, próximo àquele encontrado na varredura de genoma, que, por sua vez, foi um estudo de ligação. Interessantemente, as diferenças entre as distribuições das freqüências alélicas se deu somente no estrato europeu, indicando que a origem étnica de fato pode influenciar no desfecho da infecção por L. infantum. 60
65 Tabela 9: Análise de significância de associação entre marcadores de microssatélite e desfecho da infecção em indivíduos agrupados por origem étnica. LV x DTH+ Teste exato de Fisher LV x DTH- Marcador População geral Africano Europeu População total Africano Europeu D9S1879 0,149 0,316 0,588 0,391 0,238 0,338 D9S925 0,059 0,657 0,041* 0,222 0,502 0,033* D9S1121 0,682 0,963 0,631 0,642 0,642 0,422 D9S1118 0,610 0,311 0,060 0,626 0,347 0,679 D9S301 0,316 0,330 0,170 0,666 0,341 0,944 D9S1122 0,784 0,684 0,840 0,641 0,476 0,680 D9S922 0,675 0,380 0,813 0,854 0,921 0,797 D15S655 0,703 0,360 0,961 0,447 0,545 0,188 D15S652 0,186 0,419 0,296 0,569 0,745 0,418 D15S657 0,488 0,195 0,710 0,735 0,701 0,959 D15S966 0,370 0,361 0,676 0,701 0,409 0,829 D15S642 0,287 0,584 0,422 0,807 0,570 0,884 D19S433 0,789 0,581 0,420 0,922 0,912 0,931 D19S245 0,781 0,450 0,623 0,257 0,956 0,271 D19S178 0,649 0,304 0,891 0,873 0,345 0,948 D19S246 0,414 0,246 0,297 0,893 0,434 0,898 D19S589 0,438 0,528 0,835 0,568 0,490 0,849 D19S254 0,836 0,899 0,697 0,907 0,286 0,903 D22S692 0,265 0,566 0,521 0,502 0,601 0,874 D22S445 0,604 0,576 0,267 0,589 0,657 0,282 D22S1169 0,361 0,373 0,078 0,163 0,148 0,192 * Associação significativa 61
66 Tabela 10: Freqüências alélicas relativas do marcador D9S925. Alelos VL x DTH+ total Africano Europeu VL DTH+ VL DTH+ VL DTH+ 1* VL x DTH-ACtotal Africano Europeu VL DTH+ VL DTH+ VL DTH+ 1* * O 1 representa o conjunto dos alelos 174, 178 e 182, que apresentaram cada um freqüências menores que 5% na população. 62
67 5. DISCUSSÃO GERAL: A infecção por Leishmania pode evoluir de forma crônica e, mesmo quando tratada, pode persistir em células do sistema imunológico, principalmente macrófagos e células dendríticas ( ). Dados mostram que provalvemente não há cura parasitológica, mesmo com tratamento específico. A infecção por Leishmania infantum pode evoluir para doença naqueles indivíduos que apresentam falha em iniciar a resposta Th1. É então estabelecido um estado no qual a evolução na interrelação entre Leishmaniahospedeiro depende de mecanismos de escape que as diferentes espécies de Leishmania apresentam e a capacidade de resposta do sistema imunológico do hospedeiro, capacidade essa que varia e acordo com fatores ambientais e genéticos. Evidentemente que há fatores ambientais confundidores, como a falta de condições de saneamento e carência nutricional, fatores relativos aos vetores, que podem modular a resposta imune, mas os estudos de varredura apontam diferenças genéticas marcantes (213;214). Determinadas populações humanas se originaram a partir de pequenas quantidades de pessoas que migraram de um local a outro, onde se estabeleceram e expandiram. Essas populações representam um subconjunto da população original, com freqüências alélicas e genotípicas que podem ser diferentes. Esse fenômeno é denominado efeito do fundador (132). Migrações exploratórias a grandes distâncias, conflitos armados ou modificações climáticas abruptas podem levar populações a se espalhar em grupos em busca de locais geográficos com condições favoráveis ao assentamento, criando populações geradas pelo efeito do fundador. A migração e estabelecimento de índios em direção sul a partir do Alasca até a América do 63
68 sul são exemplos. Outro exemplo é o que se deu no Sudão, ano de 1984 quando, devido a uma grande seca, dezenas de indivíduos da etnia Masalit migraram e se estabeleceram na vila de Um Salala que, por sua vez, havia sido estabelecida somente em Em 1991 a vila contava com 1430 habitantes vivendo em más condições de infra-estrutura (215). No Sudão várias vilas surgiram como a de Um Salala referida acima. Tais populações comportam-se da mesma maneira com respeito à susceptibilidade à infecção por L. donovani, mas possuem diferenças genéticas interessantes. Essa variação e seus efeitos sobre ao desfecho da infecção por Leishmania foi investigada em estudos de varredura de genoma (213;214). Em 2003, Bucheton e colaboradores estudaram a população da vila de Barbar El Fugara, no leste do Sudão, que sofreu uma epidemia de leishmaniose visceral entre 1995 e A maioria das pessoas da vila pertence à etnia Aringa e se estabeleceram na região por volta de O estudo encontrou diferenças no perfil epidemiológico de indivíduos pertencentes aos diferentes grupos étnicos encontrados na vila: dos indivíduos com menos de 30 anos de idade, 34% daqueles da etnia Aringa desenvolveram a doença, contra 10% daqueles da etnia Hausa e 14% da etnia Fellata. Estudando somente os indivíduos Aringa, mais susceptíveis, o grupo encontrou uma área de associação no cromossomo 22q12, com alto LOD escore (3,50). Em 2007 duas vilas sudanesas, Um Salala e El Rugab também no leste do país, foram investigadas (214). Ambas encontram-se dentro da área endêmica para leishmaniose visceral e são povoadas por membros da etnia Masalit que povoaram a região por volta dos anos 1980, conforme citado 64
69 anteriormente. As duas vilas encontram-se a 40km uma da outra e, em 1997 contavam com 1225 (Um Salala) e 2084 (El Rugab) habitantes. Além de realizar a varredura de genoma com 360 marcadores microssatélites, Myler e colaboradores genotiparam pessoaspara marcadores de cromossomo Y. Assim como os indivíduos da etnia Aringa, os da etnia Masalit são susceptíveis à leishmaniose visceral. Este estudo encontrou picos de associação nos cromossomos 1, 5, 6 e 13, com LOD escores variando entre 0,77 a 2,83. Após estratificação da análise por vila e refinamento da varredura, foram encontrados picos de associação diferentes no cromossomo 1 para cada vila (1p22 em Um Salala e 1q31.3 em El Rugab) e um pico comum para ambas no cromossomo 6 (6q27). A estratificação por haplogrupos de cromossomo Y mostrou dois haplótipos principais em ambas as vilas, E3b1 (denominado E1b1b1a desde 2008 (168)) e A3b2. O pico de associação no cromossomo 1p22 encontrado em Um Salala teve contribuição somente dos indivíduos do haplogrupo E1b1b1a e nenhum dos indivíduos desses dois grupos mostrou associação com 1p22 em El Rugab. O pico de 6q27 nos indivíduos de El Rugab foi específico para indivíduos do haplótipo E1b1b1a e nenhum indivíduo de Um Salala, E1b1b1a ou A3b2 contribuiu para esse pico em 6q27. Apesar de os grupos étnicos serem semelhantes em termo de susceptibilidade não houve sobreposição de regiões de ligação entre os dois estudos realizados. Miller e colaboradores propõem que essa diferença pode ter ocorrido porque o efeito do fundador pode ter levado genes diferentes a controlar a susceptibilidade nas diferentes vilas. Esses resultados tomados em conjunto reforçam a idéia de que genes diferentes controlam a susceptibilidade 65
70 à leishmaniose visceral de maneira origem-específica, como conseqüência da influência do efeito do fundador. O resultado de varredura do genoma realizado em população do estado do Rio Grande do Norte, apresentado aqui, realizado, permite reforçar essa idéia de que o efeito do fundador pode exercer influência na susceptibilidade. Embora a doença no Brasil seja causada por uma espécie de Leishmania diferente, as características fisiopatológicas da LV são semelhantes. Ainda assim encontramos regiões diferentes daquelas dos estudos no Sudão (regiões de associação nos cromossomos 9, 15 e 19); Obviamente nossa população é bastante diversa daquela analisada nos estudos descritos acima, já que aquela população é geneticamente mais heterogênea enquanto que a nossa é composta por indivíduos de vários grupos étnicos. Um passo importante para a melhor avaliação das causas na discrepância entre regiões genômicas assinaladas pelo estudo de varredura seria a estratificação dos indivíduos analisados por origem étnica, utilizando marcadores de cromossomo Y como realizado por Miller e colaboradores. Foi também realizada uma varredura de genoma utilizando famílias residentes em área endêmica para leishmaniose visceral em Belém do Pará. Este estudo indicou três regiões genômicas em ligação com LV: 6q27, 7q11.22 e 17q11.2- q21.3 (216). Diferentes estudos apresentam diferentes regiões do genoma relacionadas ao desfecho da infecção por L. infantum. Tomados em conjunto esses resultados demonstram a complexidade do componente genético da resposta imune frente à infecção por L. infantum. A análise de correlação entre desfecho da infecção e background genético foi feita utilizando o Teste exato de Fisher com marcadores escolhidos 66
71 dentro a região indicada por uma varredura por ligação em todo o genoma (análise de ligação genética) previamente realizada em famílias da população de Natal (175). Os resultados revelaram que um marcador no cromossomo 9 (D9S925) está associado ao desfecho da infecção nos indivíduos de origem paterna europeia. Este resultado é interessante porque, apesar de termos realizado uma análise de associação utilizando alelos de microssatélites, conseguimos ainda uma significância em um dos marcadores. O ideal teria sido utilizar marcadores bialélicos que viabilizassem uma análise adicional de regressão logística. Ainda assim, a confirmação dessa região no cromossomo 9 nos dá a idéia de que os genes na vizinhança deste marcador devem ser avaliados em busca de variações que possam explicar a associação. Outro estudo realizado com o mesmo conjunto de indivíduos de Natal RN encontrou evidências de três subpopulações formando a estrutura étnica da população, desta vez utilizando marcadores microssatélite (176). Foi demonstrado que um marcador no cromossomo 22 (D22S1169) apresentou excesso de ancestralidade européia em indivíduos com LV. Isto pode significar que europeus que vieram explorar e colonizar a região conhecida hoje como o estado do Rio Grande do Norte carregaram alelos controlando a susceptibilidade à doença visceral causada por L. infantum. O nosso estudo não reproduziu este resultado, mas o teste Exato de Fisher que realizamos em função da origem étnica mostrou um valor p próximo da significância exatamente na comparação de indivíduos LV x DTH+ de origem européia. 67
72 6. Conclusões 1. Três regiões do genoma estão em ligação genética com os fenótipos susceptibilidade (9p21) e resistência (15q26 e 19q13.3) à LV; 2. As populações estudadas possuem origem paterna predominantemente européia, mas sua variabilidade deve-se ao componente africano; 3. Há evidências de que o cromossomo 9p22 apresenta genes associados à LV principalmente em indivíduos de origem européia, o que indica que a origem étnica de um indivíduo, ou seja, a origem de seu conjunto gênico pode influenciar no desfecho da infecção por L. infantum. 68
73 7. Referências bibliográficas (1) Costa CH, Pereira HF, Araujo MV. [Visceral leishmaniasis epidemic in the State of Piaui, Brazil, ]. Rev Saude Publica 1990 Oct;24(5): (2) Jeronimo SM, Oliveira RM, Mackay S, Costa RM, Sweet J, Nascimento ET, et al. An urban outbreak of visceral leishmaniasis in Natal, Brazil. Trans R Soc Trop Med Hyg 1994 Jul;88(4): (3) Jeronimo SM, Duggal P, Braz RF, Cheng C, Monteiro GR, Nascimento ET, et al. An emerging peri-urban pattern of infection with Leishmania chagasi, the protozoan causing visceral leishmaniasis in northeast Brazil. Scand J Infect Dis 2004;36(6-7): (4) Werneck GL, Costa CH, Walker AM, David JR, Wand M, Maguire JH. The urban spread of visceral leishmaniasis: clues from spatial analysis. Epidemiology 2002 May;13(3): (5) Murray HW, Nathan CF. Macrophage microbicidal mechanisms in vivo: reactive nitrogen versus oxygen intermediates in the killing of intracellular visceral Leishmania donovani. J Exp Med 1999 Feb 15;189(4): (6) Badaro R, Jones TC, Lorenco R, Cerf BJ, Sampaio D, Carvalho EM, et al. A prospective study of visceral leishmaniasis in an endemic area of Brazil. J Infect Dis 1986 Oct;154(4): (7) Kaye PM. The Immunology of Visceral leishmaniasis: Current status. In: Farrel JP, editor. Leishmania.Boston: Kluwer Academic Publishers; p (8) Wilson ME, Young BM, Davidson BL, Mente KA, McGowan SE. The importance of TGF-beta in murine visceral leishmaniasis. J Immunol 1998 Dec 1;161(11): (9) Badaro R, Jones TC, Lorenco R, Cerf BJ, Sampaio D, Carvalho EM, et al. A prospective study of visceral leishmaniasis in an endemic area of Brazil. J Infect Dis 1986 Oct;154(4): (10) Badaro R, Carvalho EM, Rocha H, Queiroz AC, Jones TC. Leishmania donovani: an opportunistic microbe associated with progressive disease in three immunocompromised patients. Lancet 1986 Mar 22;1(8482): (11) Jeronimo SM, Teixeira MJ, Sousa A, Thielking P, Pearson RD, Evans TG. Natural history of Leishmania (Leishmania) chagasi infection in Northeastern Brazil: long-term follow-up. Clin Infect Dis 2000 Mar;30(3): (12) Lima ID, Queiroz JW, Lacerda HG, Queiroz PV, Pontes NN, Barbosa JD, et al. Leishmania infantum chagasi in northeastern Brazil: 69
74 asymptomatic infection at the urban perimeter. Am J Trop Med Hyg 2012 Jan;86(1): (13) Nascimento ET, Moura ML, Queiroz JW, Barroso AW, Araujo AF, Rego EF, et al. The emergence of concurrent HIV-1/AIDS and visceral leishmaniasis in Northeast Brazil. Trans R Soc Trop Med Hyg 2011 May;105(5): (14) Reed SG, Badaro R, Masur H, Carvalho EM, Lorenco R, Lisboa A, et al. Selection of a skin test antigen for American visceral leishmaniasis. Am J Trop Med Hyg 1986 Jan;35(1): (15) Jeronimo SM, Teixeira MJ, Sousa A, Thielking P, Pearson RD, Evans TG. Natural history of Leishmania (Leishmania) chagasi infection in Northeastern Brazil: long-term follow-up. Clin Infect Dis 2000 Mar;30(3): (16) Evans TG, Teixeira MJ, McAuliffe IT, Vasconcelos I, Vasconcelos AW, Sousa AA, et al. Epidemiology of visceral leishmaniasis in northeast Brazil. J Infect Dis 1992 Nov;166(5): (17) Jeronimo SM, Teixeira MJ, Sousa A, Thielking P, Pearson RD, Evans TG. Natural history of Leishmania (Leishmania) chagasi infection in Northeastern Brazil: long-term follow-up. Clin Infect Dis 2000 Mar;30(3): (18) Desjeux P. Leishmaniasis: current situation and new perspectives. Comp Immunol Microbiol Infect Dis 2004 Sep;27(5): (19) Herwaldt BL. Leishmaniasis. Lancet 1999 Oct 2;354(9185): (20) Jeronimo J, Souza AQ, Pearson RD. Leishmaniasis. In: Goldman L, Ausielo D, editors. Cecil Textbook of Medicine. 22 ed. Phyladelphia, Pennsylvania: Saunders; p (21) Pearson RD, Lareau SM, Jeronimo SM. Leishmaniasis at the End of the Millennium. Curr Infect Dis Rep 1999 Dec;1(5): (22) Jeronimo SM, Oliveira RM, Mackay S, Costa RM, Sweet J, Nascimento ET, et al. An urban outbreak of visceral leishmaniasis in Natal, Brazil. Trans R Soc Trop Med Hyg 1994 Jul;88(4): (23) Haldar JP, Ghose S, Saha KC, Ghose AC. Cell-mediated immune response in Indian kala-azar and post-kala-azar dermal leishmaniasis. Infect Immun 1983 Nov;42(2): (24) Zijlstra EE, el-hassan AM. Leishmaniasis in Sudan. Visceral leishmaniasis. Trans R Soc Trop Med Hyg 2001 Apr;95 Suppl 1:S27- S58. 70
75 (25) Mauricio IL, Howard MK, Stothard JR, Miles MA. Genomic diversity in the Leishmania donovani complex. Parasitology 1999 Sep;119 ( Pt 3): (26) Aleixo JA, Nascimento ET, Monteiro GR, Fernandes MZ, Ramos AM, Wilson ME, et al. Atypical American visceral leishmaniasis caused by disseminated Leishmania amazonensis infection presenting with hepatitis and adenopathy. Trans R Soc Trop Med Hyg 2006 Jan;100(1): (27) Barral A, Pedral-Sampaio D, Grimaldi JG, Momen H, Mahon-Pratt D, Ribeiro de JA, et al. Leishmaniasis in Bahia, Brazil: evidence that Leishmania amazonensis produces a wide spectrum of clinical disease. Am J Trop Med Hyg 1991 May;44(5): (28) Alborzi A, Rasouli M, Shamsizadeh A. Leishmania tropica-isolated patient with visceral leishmaniasis in southern Iran. Am J Trop Med Hyg 2006 Feb;74(2): (29) Alborzi A, Pouladfar GR, Fakhar M, Motazedian MH, Hatam GR, Kadivar MR. Isolation of Leishmania tropica from a patient with visceral leishmaniasis and disseminated cutaneous leishmaniasis, southern Iran. Am J Trop Med Hyg 2008 Sep;79(3): (30) Britto C, Ravel C, Bastien P, Blaineau C, Pages M, Dedet JP, et al. Conserved linkage groups associated with large-scale chromosomal rearrangements between Old World and New World Leishmania genomes. Gene 1998 Nov 5;222(1): (31) Ivens AC, Peacock CS, Worthey EA, Murphy L, Aggarwal G, Berriman M, et al. The genome of the kinetoplastid parasite, Leishmania major. Science 2005 Jul 15;309(5733): (32) Ivens AC, Peacock CS, Worthey EA, Murphy L, Aggarwal G, Berriman M, et al. The genome of the kinetoplastid parasite, Leishmania major. Science 2005 Jul 15;309(5733): (33) Titus RG, Ribeiro JM. Salivary gland lysates from the sand fly Lutzomyia longipalpis enhance Leishmania infectivity. Science 1988 Mar 11;239(4845): (34) Pimenta PF, Saraiva EM, Rowton E, Modi GB, Garraway LA, Beverley SM, et al. Evidence that the vectorial competence of phlebotomine sand flies for different species of Leishmania is controlled by structural polymorphisms in the surface lipophosphoglycan. Proc Natl Acad Sci U S A 1994 Sep 13;91(19): (35) Ximenes MF, Castellon EG, de Souza MF, Freitas RA, Pearson RD, Wilson ME, et al. Distribution of phlebotomine sand flies (Diptera: Psychodidae) in the state of Rio Grande do Norte, Brazil. J Med Entomol 2000 Jan;37(1):
76 (36) Evans TG, Smith D, Pearson RD. Humoral factors and nonspecific immune suppression in Syrian hamsters infected with Leishmania donovani. J Parasitol 1990 Apr;76(2): (37) Ibrahim ME, Lambson B, Yousif AO, Deifalla NS, Alnaiem DA, Ismail A, et al. Kala-azar in a high transmission focus: an ethnic and geographic dimension. Am J Trop Med Hyg 1999 Dec;61(6): (38) Alexander B, Ferro C, Young DG, Morales A, Tesh RB. Ecology of phlebotomine sand flies (Diptera: Psychodidae) in a focus of Leishmania (Viannia) braziliensis in northeastern Colombia. Mem Inst Oswaldo Cruz 1992 Jul;87(3): (39) Jeronimo SM, Pearson RD. The Leishmania. Protozoans adapted for extracellular and intracellular survival. Subcell Biochem 1992;18:1-37. (40) Lainson R, Ward RD, Shaw JJ. Experimental transmission of Leishmania chagasi, causative agent of neotropical visceral leishmaniasis, by the sandfly Lutzomyia longipalpis. Nature 1977 Apr 14;266(5603): (41) Hall LR, Titus RG. Sand fly vector saliva selectively modulates macrophage functions that inhibit killing of Leishmania major and nitric oxide production. J Immunol 1995 Oct 1;155(7): (42) Belkaid Y, Mendez S, Lira R, Kadambi N, Milon G, Sacks D. A natural model of Leishmania major infection reveals a prolonged "silent" phase of parasite amplification in the skin before the onset of lesion formation and immunity. J Immunol 2000 Jul 15;165(2): (43) Collin N, Gomes R, Teixeira C, Cheng L, Laughinghouse A, Ward JM, et al. Sand fly salivary proteins induce strong cellular immunity in a natural reservoir of visceral leishmaniasis with adverse consequences for Leishmania. PLoS Pathog 2009 May;5(5):e (44) Blackwell JM. Genetic susceptibility to leishmanial infections: studies in mice and man. Parasitology 1996;112 Suppl:S67-S74. (45) Evans T, Naidu TG, de Alencar JE, Pearson RD. The relationship of American visceral leishmaniasis to ABO blood group type. Am J Trop Med Hyg 1984 Sep;33(5): (46) Evans TG, Teixeira MJ, McAuliffe IT, Vasconcelos I, Vasconcelos AW, Sousa AA, et al. Epidemiology of visceral leishmaniasis in northeast Brazil. J Infect Dis 1992 Nov;166(5): (47) Garin YJ, Sulahian A, Gangneux JP, Derouin F. Activity of IFN-gamma in experimental Leishmania infantum murine visceral leishmaniasis. Pathol Biol (Paris) 1997 Jan;45(1):
77 (48) Badaro R, Jones TC, Lorenco R, Cerf BJ, Sampaio D, Carvalho EM, et al. A prospective study of visceral leishmaniasis in an endemic area of Brazil. J Infect Dis 1986 Oct;154(4): (49) Harrison LH, Naidu TG, Drew JS, de Alencar JE, Pearson RD. Reciprocal relationships between undernutrition and the parasitic disease visceral leishmaniasis. Rev Infect Dis 1986 May;8(3): (50) Mathis A, Deplazes P. PCR and in vitro cultivation for detection of Leishmania spp. in diagnostic samples from humans and dogs. J Clin Microbiol 1995 May;33(5): (51) Schallig HD, Oskam L. Molecular biological applications in the diagnosis and control of leishmaniasis and parasite identification. Trop Med Int Health 2002 Aug;7(8): (52) Silva AV, Paula AA, Cabrera MA, Carreira JC. [Leishmaniasis in domestic dogs: epidemiological aspects]. Cad Saude Publica 2005 Jan;21(1): (53) Silva ES, Pacheco RS, Gontijo CM, Carvalho IR, Brazil RP. Visceral leishmaniasis caused by Leishmania (Viannia) braziliensis in a patient infected with human immunodeficiency virus. Rev Inst Med Trop Sao Paulo 2002 May;44(3): (54) Jeronimo SM, Oliveira RM, Mackay S, Costa RM, Sweet J, Nascimento ET, et al. An urban outbreak of visceral leishmaniasis in Natal, Brazil. Trans R Soc Trop Med Hyg 1994 Jul;88(4): (55) Boom WH, Liebster L, Abbas AK, Titus RG. Patterns of cytokine secretion in murine leishmaniasis: correlation with disease progression or resolution. Infect Immun 1990 Dec;58(12): (56) Coffman RL, Mocci S, O'Garra A. The stability and reversibility of Th1 and Th2 populations. Curr Top Microbiol Immunol 1999;238:1-12. (57) Murphy ML, Cotterell SE, Gorak PM, Engwerda CR, Kaye PM. Blockade of CTLA-4 enhances host resistance to the intracellular pathogen, Leishmania donovani. J Immunol 1998 Oct 15;161(8): (58) Kaplan MH, Sun YL, Hoey T, Grusby MJ. Impaired IL-12 responses and enhanced development of Th2 cells in Stat4-deficient mice. Nature 1996 Jul 11;382(6587): (59) Thierfelder WE, van Deursen JM, Yamamoto K, Tripp RA, Sarawar SR, Carson RT, et al. Requirement for Stat4 in interleukin-12-mediated responses of natural killer and T cells. Nature 1996 Jul 11;382(6587):
78 (60) Wilson ME, Young BM, Davidson BL, Mente KA, McGowan SE. The importance of TGF-beta in murine visceral leishmaniasis. J Immunol 1998 Dec 1;161(11): (61) Rezai HR, Farrell J, Soulsby EL. Immunological responses of L. donovani infection in mice and significance of T cell in resistance to experimental leishmaniasis. Clin Exp Immunol 1980 Jun;40(3): (62) Skov CB, Twohy DW. Cellular immunity to Leishmania donovani. I. The effect of T cell depletion on resistance to L. donovani in mice. J Immunol 1974 Dec;113(6): (63) Murray HW. Cell-mediated immune response in experimental visceral leishmaniasis. II. Oxygen-dependent killing of intracellular Leishmania donovani amastigotes. J Immunol 1982 Jul;129(1): (64) Murray HW, Masur H, Keithly JS. Cell-mediated immune response in experimental visceral leishmaniasis. I. Correlation between resistance to Leishmania donovani and lymphokine-generating capacity. J Immunol 1982 Jul;129(1): (65) Belosevic M, Finbloom DS, Meltzer MS, Nacy CA. IL-2. A cofactor for induction of activated macrophage resistance to infection. J Immunol 1990 Aug 1;145(3): (66) Kumar R, Pai K, Sundar S. Reactive oxygen intermediates, nitrite and IFN-gamma in Indian visceral leishmaniasis. Clin Exp Immunol 2001 May;124(2): (67) Liew FY. Induction and regulation of CD4+ T cell subsets. Ciba Found Symp 1994;187: (68) Nacy CA, Nelson BJ, Meltzer MS, Green SJ. Cytokines that regulate macrophage production of nitrogen oxides and expression of antileishmanial activities. Res Immunol 1991 Sep;142(7): (69) Nacy CA, Meierovics AI, Belosevic M, Green SJ. Tumor necrosis factor-alpha: central regulatory cytokine in the induction of macrophage antimicrobial activities. Pathobiology 1991;59(3): (70) Murray PJ, Handman E, Glaser TA, Spithill TW. Leishmania major: expression and gene structure of the glycoprotein 63 molecule in virulent and avirulent clones and strains. Exp Parasitol 1990 Oct;71(3): (71) Rao A, Luo C, Hogan PG. Transcription factors of the NFAT family: regulation and function. Annu Rev Immunol 1997;15: (72) Green SJ, Mellouk S, Hoffman SL, Meltzer MS, Nacy CA. Cellular mechanisms of nonspecific immunity to intracellular infection: cytokineinduced synthesis of toxic nitrogen oxides from L-arginine by macrophages and hepatocytes. Immunol Lett 1990 Aug;25(1-3):
79 (73) Green SJ, Meltzer MS, Hibbs JB, Jr., Nacy CA. Activated macrophages destroy intracellular Leishmania major amastigotes by an L-argininedependent killing mechanism. J Immunol 1990 Jan 1;144(1): (74) Murray HW. Susceptibility of Leishmania to oxygen intermediates and killing by normal macrophages. J Exp Med 1981 May 1;153(5): (75) Murray HW. Cell-mediated immune response in experimental visceral leishmaniasis. II. Oxygen-dependent killing of intracellular Leishmania donovani amastigotes. J Immunol 1982 Jul;129(1): (76) Diefenbach A, Schindler H, Donhauser N, Lorenz E, Laskay T, MacMicking J, et al. Type 1 interferon (IFNalpha/beta) and type 2 nitric oxide synthase regulate the innate immune response to a protozoan parasite. Immunity 1998 Jan;8(1): (77) Bacellar O, D'oliveira A, Jr., Jeronimo S, Carvalho EM. IL-10 and IL-12 are the main regulatory cytokines in visceral leishmaniasis. Cytokine 2000 Aug;12(8): (78) Carvalho EM, Bacellar O, Brownell C, Regis T, Coffman RL, Reed SG. Restoration of IFN-gamma production and lymphocyte proliferation in visceral leishmaniasis. J Immunol 1994 Jun 15;152(12): (79) Ghalib HW, Whittle JA, Kubin M, Hashim FA, el-hassan AM, Grabstein KH, et al. IL-12 enhances Th1-type responses in human Leishmania donovani infections. J Immunol 1995 May 1;154(9): (80) Ribeiro-de-Jesus A, Almeida RP, Lessa H, Bacellar O, Carvalho EM. Cytokine profile and pathology in human leishmaniasis. Braz J Med Biol Res 1998 Jan;31(1): (81) Miles SA, Conrad SM, Alves RG, Jeronimo SM, Mosser DM. A role for IgG immune complexes during infection with the intracellular pathogen Leishmania. J Exp Med 2005 Mar 7;201(5): (82) Evans TG, Reed SS, Hibbs JB, Jr. Nitric oxide production in murine leishmaniasis: correlation of progressive infection with increasing systemic synthesis of nitric oxide. Am J Trop Med Hyg 1996 May;54(5): (83) Roach TI, Kiderlen AF, Blackwell JM. Role of inorganic nitrogen oxides and tumor necrosis factor alpha in killing Leishmania donovani amastigotes in gamma interferon-lipopolysaccharide-activated macrophages from Lshs and Lshr congenic mouse strains. Infect Immun 1991 Nov;59(11): (84) Dereure J, El-Safi SH, Bucheton B, Boni M, Kheir MM, Davoust B, et al. Visceral leishmaniasis in eastern Sudan: parasite identification in humans and dogs; host-parasite relationships. Microbes Infect 2003 Oct;5(12):
80 (85) Ramirez JL, Guevara P. Persistent infections by Leishmania (Viannia) braziliensis. Mem Inst Oswaldo Cruz 1997 May;92(3): (86) Schubach A, Haddad F, Oliveira-Neto MP, Degrave W, Pirmez C, Grimaldi G, Jr., et al. Detection of Leishmania DNA by polymerase chain reaction in scars of treated human patients. J Infect Dis 1998 Sep;178(3): (87) Schubach A, Marzochi MC, Cuzzi-Maya T, Oliveira AV, Araujo ML, Oliveira AL, et al. Cutaneous scars in American tegumentary leishmaniasis patients: a site of Leishmania (Viannia) braziliensis persistence and viability eleven years after antimonial therapy and clinical cure. Am J Trop Med Hyg 1998 Jun;58(6): (88) Stenger S, Donhauser N, Thuring H, Rollinghoff M, Bogdan C. Reactivation of latent leishmaniasis by inhibition of inducible nitric oxide synthase. J Exp Med 1996 Apr 1;183(4): (89) Badaro R, Carvalho EM, Rocha H, Queiroz AC, Jones TC. Leishmania donovani: an opportunistic microbe associated with progressive disease in three immunocompromised patients. Lancet 1986 Mar 22;1(8482): (90) Hernandez-Perez J, Yebra-Bango M, Jimenez-Martinez E, Sanz- Moreno C, Cuervas-Mons V, Alonso PL, et al. Visceral leishmaniasis (kala-azar) in solid organ transplantation: report of five cases and review. Clin Infect Dis 1999 Oct;29(4): (91) Rihl M, Stoll M, Ulbricht K, Bange FC, Schmidt RE. Successful treatment of post-kala-azar dermal leishmaniasis (PKDL) in a HIV infected patient with multiple relapsing leishmaniasis from Western Europe. J Infect 2006 Jul;53(1):e25-e27. (92) Belkaid Y, Hoffmann KF, Mendez S, Kamhawi S, Udey MC, Wynn TA, et al. The role of interleukin (IL)-10 in the persistence of Leishmania major in the skin after healing and the therapeutic potential of anti-il-10 receptor antibody for sterile cure. J Exp Med 2001 Nov 19;194(10): (93) Belkaid Y, Piccirillo CA, Mendez S, Shevach EM, Sacks DL. CD4+CD25+ regulatory T cells control Leishmania major persistence and immunity. Nature 2002 Dec 5;420(6915): (94) Bluestone JA, Tang Q. How do CD4+CD25+ regulatory T cells control autoimmunity? Curr Opin Immunol 2005 Dec;17(6): (95) Sakaguchi S, Sakaguchi N, Asano M, Itoh M, Toda M. Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of selftolerance causes various autoimmune diseases. J Immunol 1995 Aug 1;155(3):
81 (96) Sakaguchi S, Sakaguchi N. Regulatory T cells in immunologic selftolerance and autoimmune disease. Int Rev Immunol 2005 May;24(3-4): (97) Sakaguchi S. Regulatory T cells. Springer Semin Immunopathol 2006 Aug;28(1):1-2. (98) Aebischer T, Moody SF, Handman E. Persistence of virulent Leishmania major in murine cutaneous leishmaniasis: a possible hazard for the host. Infect Immun 1993 Jan;61(1): (99) Stenger S, Thuring H, Rollinghoff M, Bogdan C. Tissue expression of inducible nitric oxide synthase is closely associated with resistance to Leishmania major. J Exp Med 1994 Sep 1;180(3): (100) Stenger S, Donhauser N, Thuring H, Rollinghoff M, Bogdan C. Reactivation of latent leishmaniasis by inhibition of inducible nitric oxide synthase. J Exp Med 1996 Apr 1;183(4): (101) Belkaid Y, Piccirillo CA, Mendez S, Shevach EM, Sacks DL. CD4+CD25+ regulatory T cells control Leishmania major persistence and immunity. Nature 2002 Dec 5;420(6915): (102) Mendez S, Reckling SK, Piccirillo CA, Sacks D, Belkaid Y. Role for CD4(+) CD25(+) regulatory T cells in reactivation of persistent leishmaniasis and control of concomitant immunity. J Exp Med 2004 Jul 19;200(2): (103) Aseffa A, Gumy A, Launois P, Macdonald HR, Louis JA, Tacchini- Cottier F. The early IL-4 response to Leishmania major and the resulting Th2 cell maturation steering progressive disease in BALB/c mice are subject to the control of regulatory CD4+CD25+ T cells. J Immunol 2002 Sep 15;169(6): (104) Belkaid Y, Piccirillo CA, Mendez S, Shevach EM, Sacks DL. CD4+CD25+ regulatory T cells control Leishmania major persistence and immunity. Nature 2002 Dec 5;420(6915): (105) Blackwell JM, Black GF, Peacock CS, Miller EN, Sibthorpe D, Gnananandha D, et al. Immunogenetics of leishmanial and mycobacterial infections: the Belem Family Study. Philos Trans R Soc Lond B Biol Sci 1997 Sep 29;352(1359): (106) Hawn TR, Ozinsky A, Underhill DM, Buckner FS, Akira S, Aderem A. Leishmania major activates IL-1 alpha expression in macrophages through a MyD88-dependent pathway. Microbes Infect 2002 Jul;4(8): (107) Newport M, Levin M. Genetic susceptibility to tuberculosis. J Infect 1999 Sep;39(2):
82 (108) Newport MJ, Huxley CM, Huston S, Hawrylowicz CM, Oostra BA, Williamson R, et al. A mutation in the interferon-gamma-receptor gene and susceptibility to mycobacterial infection. N Engl J Med 1996 Dec 26;335(26): (109) Roberds SL. Applying genomics tools to identify therapeutic targets for asthma. Expert Opin Investig Drugs 1998 Aug;7(8): (110) Stockton JC, Howson JM, Awomoyi AA, McAdam KP, Blackwell JM, Newport MJ. Polymorphism in NOD2, Crohn's disease, and susceptibility to pulmonary tuberculosis. FEMS Immunol Med Microbiol 2004 Jun 1;41(2): (111) Yee LJ. Host genetic determinants in hepatitis C virus infection. Genes Immun 2004 Jun;5(4): (112) Wang H, Mengsteab S, Tag CG, Gao CF, Hellerbrand C, Lammert F, et al. Transforming growth factor-beta1 gene polymorphisms are associated with progression of liver fibrosis in Caucasians with chronic hepatitis C infection. World J Gastroenterol 2005 Apr 7;11(13): (113) Lazarus R, Raby BA, Lange C, Silverman EK, Kwiatkowski DJ, Vercelli D, et al. TOLL-like receptor 10 genetic variation is associated with asthma in two independent samples. Am J Respir Crit Care Med 2004 Sep 15;170(6): (114) van Rietschoten JG, Westland R, van den BR, Nieste-Otter MA, van VA, Jonkers RE, et al. A novel polymorphic GATA site in the human IL- 12Rbeta2 promoter region affects transcriptional activity. Tissue Antigens 2004 Jun;63(6): (115) Fife MS, Fisher SA, John S, Worthington J, Shah CJ, Ollier WE, et al. Multipoint linkage analysis of a candidate gene locus in rheumatoid arthritis demonstrates significant evidence of linkage and association with the corticotropin-releasing hormone genomic region. Arthritis Rheum 2000 Aug;43(8): (116) Palmer LJ, Cookson WO. Using single nucleotide polymorphisms as a means to understanding the pathophysiology of asthma. Respir Res 2001;2(2): (117) Ferreira RG, Moura MM, Engracia V, Pagotto RC, Alves FP, Camargo LM, et al. Ethnic admixture composition of two western Amazonian populations. Hum Biol 2002 Aug;74(4): (118) Silva WA, Jr., Bonatto SL, Holanda AJ, Ribeiro-Dos-Santos AK, Paixao BM, Goldman GH, et al. Mitochondrial genome diversity of Native Americans supports a single early entry of founder populations into America. Am J Hum Genet 2002 Jul;71(1):
83 (119) Ferreira RG, Moura MM, Engracia V, Pagotto RC, Alves FP, Camargo LM, et al. Ethnic admixture composition of two western Amazonian populations. Hum Biol 2002 Aug;74(4): (120) Silva WA, Jr., Bonatto SL, Holanda AJ, Ribeiro-Dos-Santos AK, Paixao BM, Goldman GH, et al. Mitochondrial genome diversity of Native Americans supports a single early entry of founder populations into America. Am J Hum Genet 2002 Jul;71(1): (121) Magnoli D. Uma Gota de Sangue: História do Pensamento Racial. 1 ed. São Paulo: Contexto; (122) Moraes E. A Escravidão Africana no Brasil: das Origens à extinção. 3 ed. Brasília: (123) Salles V. O Negro no Pará. 1 ed (124) Batista dos Santos SE, Rodrigues JD, Ribeiro-Dos-Santos AK, Zago MA. Differential contribution of indigenous men and women to the formation of an urban population in the Amazon region as revealed by mtdna and Y-DNA. Am J Phys Anthropol 1999 Jun;109(2): (125) Carvalho-Silva DR, Santos FR, Rocha J, Pena SD. The phylogeography of Brazilian Y-chromosome lineages. Am J Hum Genet 2001 Jan;68(1): (126) Ribeiro-Dos-Santos AK, Pereira JM, Lobato MR, Carvalho BM, Guerreiro JF, Batista dos Santos SE. Dissimilarities in the process of formation of Curiau, a semi-isolated Afro-Brazilian population of the Amazon region. Am J Hum Biol 2002 Jul;14(4): (127) Parra FC, Amado RC, Lambertucci JR, Rocha J, Antunes CM, Pena SD. Color and genomic ancestry in Brazilians. Proc Natl Acad Sci U S A 2003 Jan 7;100(1): (128) Batista dos Santos SE, Rodrigues JD, Ribeiro-Dos-Santos AK, Zago MA. Differential contribution of indigenous men and women to the formation of an urban population in the Amazon region as revealed by mtdna and Y-DNA. Am J Phys Anthropol 1999 Jun;109(2): (129) Ribeiro-Dos-Santos AK, Pereira JM, Lobato MR, Carvalho BM, Guerreiro JF, Batista dos Santos SE. Dissimilarities in the process of formation of Curiau, a semi-isolated Afro-Brazilian population of the Amazon region. Am J Hum Biol 2002 Jul;14(4): (130) Carvalho-Silva DR, Santos FR, Rocha J, Pena SD. The phylogeography of Brazilian Y-chromosome lineages. Am J Hum Genet 2001 Jan;68(1): (131) Carvalho-Silva DR, Santos FR, Rocha J, Pena SD. The phylogeography of Brazilian Y-chromosome lineages. Am J Hum Genet 2001 Jan;68(1):
84 (132) Ridley M. Evolution. 3 ed. Blackwell; (133) Parra FC, Amado RC, Lambertucci JR, Rocha J, Antunes CM, Pena SD. Color and genomic ancestry in Brazilians. Proc Natl Acad Sci U S A 2003 Jan 7;100(1): (134) Rosenzweig DY, Stead WW. The role of tuberculosis and other forms of bronchopulmonary necrosis in the pathogenesis of bronchiectasis. Am Rev Respir Dis 1966 May;93(5): (135) Rosenzweig SD, Dorman SE, Uzel G, Shaw S, Scurlock A, Brown MR, et al. A novel mutation in IFN-gamma receptor 2 with dominant negative activity: biological consequences of homozygous and heterozygous states. J Immunol 2004 Sep 15;173(6): (136) Tso HW, Lau YL, Tam CM, Wong HS, Chiang AK. Associations between IL12B polymorphisms and tuberculosis in the Hong Kong Chinese population. J Infect Dis 2004 Sep 1;190(5): (137) Ahmad T, Zhang L, Gogus F, Verity D, Wallace G, Madanat W, et al. CARD15 polymorphisms in Behcet's disease. Scand J Rheumatol 2005 May;34(3): (138) Hu DN, Yu GP, McCormick SA, Schneider S, Finger PT. Populationbased incidence of uveal melanoma in various races and ethnic groups. Am J Ophthalmol 2005 Oct;140(4): (139) Tso HW, Lau YL, Tam CM, Wong HS, Chiang AK. Associations between IL12B polymorphisms and tuberculosis in the Hong Kong Chinese population. J Infect Dis 2004 Sep 1;190(5): (140) Ibrahim ME, Lambson B, Yousif AO, Deifalla NS, Alnaiem DA, Ismail A, et al. Kala-azar in a high transmission focus: an ethnic and geographic dimension. Am J Trop Med Hyg 1999 Dec;61(6): (141) Goswami T, Bhattacharjee A, Babal P, Searle S, Moore E, Li M, et al. Natural-resistance-associated macrophage protein 1 is an H+/bivalent cation antiporter. Biochem J 2001 Mar 15;354(Pt 3): (142) Blackwell JM, Mohamed HS, Ibrahim ME. Genetics and visceral leishmaniasis in the Sudan: seeking a link. Trends Parasitol 2004 Jun;20(6): (143) Mohamed HS, Ibrahim ME, Miller EN, White JK, Cordell HJ, Howson JM, et al. SLC11A1 (formerly NRAMP1) and susceptibility to visceral leishmaniasis in The Sudan. Eur J Hum Genet 2004 Jan;12(1): (144) Miller EN, Fadl M, Mohamed HS, Elzein A, Jamieson SE, Cordell HJ, et al. Y chromosome lineage- and village-specific genes on chromosomes 1p22 and 6q27 control visceral leishmaniasis in Sudan. PLoS Genet 2007 May 11;3(5):e71. 80
85 (145) Miller EN, Fadl M, Mohamed HS, Elzein A, Jamieson SE, Cordell HJ, et al. Y chromosome lineage- and village-specific genes on chromosomes 1p22 and 6q27 control visceral leishmaniasis in Sudan. PLoS Genet 2007 May 11;3(5):e71. (146) Jeronimo SM, Oliveira RM, Mackay S, Costa RM, Sweet J, Nascimento ET, et al. An urban outbreak of visceral leishmaniasis in Natal, Brazil. Trans R Soc Trop Med Hyg 1994 Jul;88(4): (147) Karplus TM, Jeronimo SM, Chang H, Helms BK, Burns TL, Murray JC, et al. Association between the tumor necrosis factor locus and the clinical outcome of Leishmania chagasi infection. Infect Immun 2002 Dec;70(12): (148) Braz RF, Nascimento ET, Martins DR, Wilson ME, Pearson RD, Reed SG, et al. The sensitivity and specificity of Leishmania chagasi recombinant K39 antigen in the diagnosis of American visceral leishmaniasis and in differentiating active from subclinical infection. Am J Trop Med Hyg 2002 Oct;67(4): (149) Evans TG, Teixeira MJ, McAuliffe IT, Vasconcelos I, Vasconcelos AW, Sousa AA, et al. Epidemiology of visceral leishmaniasis in northeast Brazil. J Infect Dis 1992 Nov;166(5): (150) Reed SG, Badaro R, Masur H, Carvalho EM, Lorenco R, Lisboa A, et al. Selection of a skin test antigen for American visceral leishmaniasis. Am J Trop Med Hyg 1986 Jan;35(1): (151) Jeronimo SM, Duggal P, Braz RF, Cheng C, Monteiro GR, Nascimento ET, et al. An emerging peri-urban pattern of infection with Leishmania chagasi, the protozoan causing visceral leishmaniasis in northeast Brazil. Scand J Infect Dis 2004;36(6-7): (152) Jose FF, da S, I, Araujo MI, Almeida RP, Bacellar O, Carvalho EM. [Evaluation of the sensitization power of Montenegro skin test]. Rev Soc Bras Med Trop 2001 Nov;34(6): (153) Braz RF, Nascimento ET, Martins DR, Wilson ME, Pearson RD, Reed SG, et al. The sensitivity and specificity of Leishmania chagasi recombinant K39 antigen in the diagnosis of American visceral leishmaniasis and in differentiating active from subclinical infection. Am J Trop Med Hyg 2002 Oct;67(4): (154) Evans TG, Teixeira MJ, McAuliffe IT, Vasconcelos I, Vasconcelos AW, Sousa AA, et al. Epidemiology of visceral leishmaniasis in northeast Brazil. J Infect Dis 1992 Nov;166(5): (155) Evans TG, Teixeira MJ, Sousa AQ, Pearson RD. Short report: extended follow-up of the natural history of persons infected with Leishmania chagasi. Am J Trop Med Hyg 1995 Oct;53(4):
86 (156) Jeronimo SM, Teixeira MJ, Sousa A, Thielking P, Pearson RD, Evans TG. Natural history of Leishmania (Leishmania) chagasi infection in Northeastern Brazil: long-term follow-up. Clin Infect Dis 2000 Mar;30(3): (157) Ulczak OM, Ghadirian E, Skamene E, Blackwell JM, Kongshavn PA. Characterization of protective T cells in the acquired response to Leishmania donovani in genetically determined cure (H-2b) and noncure (H-2d) mouse strains. Infect Immun 1989 Sep;57(9): (158) Abecasis GR, Cherny SS, Cookson WO, Cardon LR. Merlin - Rapid analysis of dense genetic maps using sparse gene flow trees. Nature Genet. 30, Ref Type: Journal (Full) (159) Amos CI. Robust variance-components approach for assessing genetic linkage in pedigrees. Am J Hum Genet 1994 Mar;54(3): (160) Whittemore AS, Halpern J. Probability of gene identity by descent: computation and applications. Biometrics 1994 Mar;50(1): (161) Kong A, Cox NJ. Allele-sharing models: LOD scores and accurate linkage tests. Am J Hum Genet 1997 Nov;61(5): (162) Sawcer S, Jones HB, Judge D, Visser F, Compston A, Goodfellow PN, et al. Empirical genomewide significance levels established by whole genome simulations. Genet Epidemiol 1997;14(3): (163) Blackwell JM, Black GF, Peacock CS, Miller EN, Sibthorpe D, Gnananandha D, et al. Immunogenetics of leishmanial and mycobacterial infections: the Belem Family Study. Philos Trans R Soc Lond B Biol Sci 1997 Sep 29;352(1359): (164) Castellucci L, Cheng LH, Araujo C, Guimaraes LH, Lessa H, Machado P, et al. Familial aggregation of mucosal leishmaniasis in northeast Brazil. Am J Trop Med Hyg 2005 Jul;73(1): (165) Goncalves R, Freitas A, Branco M, Rosa A, Fernandes AT, Zhivotovsky LA, et al. Y-chromosome lineages from Portugal, Madeira and Acores record elements of Sephardim and Berber ancestry. Ann Hum Genet 2005 Jul;69(Pt 4): (166) Underhill PA, Passarino G, Lin AA, Shen P, Mirazon LM, Foley RA, et al. The phylogeography of Y chromosome binary haplotypes and the origins of modern human populations. Ann Hum Genet 2001 Jan;65(Pt 1): (167) Underhill PA, Passarino G, Lin AA, Shen P, Mirazon LM, Foley RA, et al. The phylogeography of Y chromosome binary haplotypes and the origins of modern human populations. Ann Hum Genet 2001 Jan;65(Pt 1):
87 (168) Karafet TM, Mendez FL, Meilerman MB, Underhill PA, Zegura SL, Hammer MF. New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree. Genome Res 2008 May;18(5): (169) A nomenclature system for the tree of human Y-chromosomal binary haplogroups. Genome Res 2002 Feb;12(2): (170) Karafet TM, Mendez FL, Meilerman MB, Underhill PA, Zegura SL, Hammer MF. New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree. Genome Res 2008 May;18(5): (171) Excoffier L, Heckel G. Computer programs for population genetics data analysis: a survival guide. Nat Rev Genet 2006 Oct;7(10): (172) Cockerham CC, Weir BS. Covariances of relatives stemming from a population undergoing mixed self and random mating. Biometrics 1984 Mar;40(1): (173) Excoffier L, Smouse PE, Quattro JM. Analysis of molecular variance inferred from metric distances among DNA haplotypes: application to human mitochondrial DNA restriction data. Genetics 1992 Jun;131(2): (174) Reynolds J, Weir BS, Cockerham CC. Estimation of the Coancestry Coefficient: Basis for a Short-Term Genetic Distance. Genetics 1983 Nov;105(3): (175) Jeronimo SM, Duggal P, Ettinger NA, Nascimento ET, Monteiro GR, Cabral AP, et al. Genetic predisposition to self-curing infection with the protozoan Leishmania chagasi: a genomewide scan. J Infect Dis 2007 Oct 15;196(8): (176) Ettinger NA, Duggal P, Braz RF, Nascimento ET, Beaty TH, Jeronimo SM, et al. Genetic admixture in Brazilians exposed to infection with Leishmania chagasi. Ann Hum Genet 2009 May;73(Pt 3): (177) Wigginton JE, Abecasis GR. PEDSTATS: descriptive statistics, graphics and quality assessment for gene mapping data. Bioinformatics 2005 Aug 15;21(16): (178) Jeronimo SM, Oliveira RM, Mackay S, Costa RM, Sweet J, Nascimento ET, et al. An urban outbreak of visceral leishmaniasis in Natal, Brazil. Trans R Soc Trop Med Hyg 1994 Jul;88(4): (179) Braz RF, Nascimento ET, Martins DR, Wilson ME, Pearson RD, Reed SG, et al. The sensitivity and specificity of Leishmania chagasi recombinant K39 antigen in the diagnosis of American visceral leishmaniasis and in differentiating active from subclinical infection. Am J Trop Med Hyg 2002 Oct;67(4):
88 (180) Reed SG, Badaro R, Masur H, Carvalho EM, Lorenco R, Lisboa A, et al. Selection of a skin test antigen for American visceral leishmaniasis. Am J Trop Med Hyg 1986 Jan;35(1): (181) Ali A, Ashford RW. Visceral leishmaniasis in Ethiopia. II. Annual leishmanin transformation in a population. Is positive leishmanin reaction a life-long phenomenon? Ann Trop Med Parasitol 1993 Apr;87(2): (182) Lander E, Kruglyak L. Genetic dissection of complex traits: guidelines for interpreting and reporting linkage results. Nat Genet 1995 Nov;11(3): (183) Bucheton B, Abel L, El-Safi S, Kheir MM, Pavek S, Lemainque A, et al. A major susceptibility locus on chromosome 22q12 plays a critical role in the control of kala-azar. Am J Hum Genet 2003 Nov;73(5): (184) Mohamed HS, Ibrahim ME, Miller EN, Peacock CS, Khalil EA, Cordell HJ, et al. Genetic susceptibility to visceral leishmaniasis in The Sudan: linkage and association with IL4 and IFNGR1. Genes Immun 2003 Jul;4(5): (185) Jamieson SE, Miller EN, Peacock CS, Fakiola M, Wilson ME, Bales- Holst A, et al. Genome-wide scan for visceral leishmaniasis susceptibility genes in Brazil. Genes Immun 2007 Jan;8(1): (186) Abe-Sandes K, Silva WA Jr, Zago MA. Heterogeneity of the Y chromosome in Afro-Brazilian populations. Hum.Biol. 76[1], Ref Type: Journal (Full) (187) Kohlrausch FB, Callegari-Jacques SM, Tsuneto LT, Ptzl-Herler ML, Hill K, Hurtado AM, et al. Geography influences microsatellite polymorphism diversity in Amerindians. Am.J.Phys Anthropol 126[4], Ref Type: Journal (Full) (188) Karafet TM, Mendez FL, Meilerman MB, Underhill PA, Zegura SL, Hammer MF. New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree. Genome Res 2008 May;18(5): (189) Su B, Jin L, Underhill P, Martinson J, Saha N, McGarvey ST, et al. Polynesian origins: insights from the Y chromosome. Proc Natl Acad Sci U S A 2000 Jul 18;97(15): (190) Underhill PA, Passarino G, Lin AA, Shen P, Mirazon LM, Foley RA, et al. The phylogeography of Y chromosome binary haplotypes and the origins of modern human populations. Ann Hum Genet 2001 Jan;65(Pt 1):
89 (191) Batista dos Santos SE, Rodrigues JD, Ribeiro-Dos-Santos AK, Zago MA. Differential contribution of indigenous men and women to the formation of an urban population in the Amazon region as revealed by mtdna and Y-DNA. Am J Phys Anthropol 1999 Jun;109(2): (192) Ribeiro-Dos-Santos AK, Pereira JM, Lobato MR, Carvalho BM, Guerreiro JF, Batista dos Santos SE. Dissimilarities in the process of formation of Curiau, a semi-isolated Afro-Brazilian population of the Amazon region. Am J Hum Biol 2002 Jul;14(4): (193) Rosser ZH, Zerjal T, Hurles ME, Adojaan M, Alavantic D, Amorim A, et al. Y-chromosomal diversity in Europe is clinal and influenced primarily by geography, rather than by language. Am J Hum Genet 2000 Dec;67(6): (194) Nebel A, Filon D, Brinkmann B, Majumder PP, Faerman M, Oppenheim A. The Y chromosome pool of Jews as part of the genetic landscape of the Middle East. Am J Hum Genet 2001 Nov;69(5): (195) Nebel A, Filon D, Faerman M, Soodyall H, Oppenheim A. Y chromosome evidence for a founder effect in Ashkenazi Jews. Eur J Hum Genet 2005 Mar;13(3): (196) Sutton WK, Knight A, Underhill PA, Neulander JS, Disotell TR, Mountain JL. Toward resolution of the debate regarding purported crypto-jews in a Spanish-American population: evidence from the Y chromosome. Ann Hum Biol 2006 Jan;33(1): (197) Behar DM, Thomas MG, Skorecki K, Hammer MF, Bulygina E, Rosengarten D, et al. Multiple origins of Ashkenazi Levites: Y chromosome evidence for both Near Eastern and European ancestries. Am J Hum Genet 2003 Oct;73(4): (198) Adams SM, Bosch E, Balaresque PL, Ballereau SJ, Lee AC, Arroyo E, et al. The genetic legacy of religious diversity and intolerance: paternal lineages of Christians, Jews, and Muslims in the Iberian Peninsula. Am J Hum Genet 2008 Dec;83(6): (199) Goncalves R, Freitas A, Branco M, Rosa A, Fernandes AT, Zhivotovsky LA, et al. Y-chromosome lineages from Portugal, Madeira and Acores record elements of Sephardim and Berber ancestry. Ann Hum Genet 2005 Jul;69(Pt 4): (200) Sutton WK, Knight A, Underhill PA, Neulander JS, Disotell TR, Mountain JL. Toward resolution of the debate regarding purported crypto-jews in a Spanish-American population: evidence from the Y chromosome. Ann Hum Biol 2006 Jan;33(1): (201) Semino O, Magri C, Benuzzi G, Lin AA, Al-Zahery N, Battaglia V, et al. Origin, diffusion, and differentiation of Y-chromosome haplogroups E and J: inferences on the neolithization of Europe and later migratory 85
90 events in the Mediterranean area. Am J Hum Genet 2004 May;74(5): (202) Underhill PA, Passarino G, Lin AA, Shen P, Mirazon LM, Foley RA, et al. The phylogeography of Y chromosome binary haplotypes and the origins of modern human populations. Ann Hum Genet 2001 Jan;65(Pt 1): (203) Semino O, Magri C, Benuzzi G, Lin AA, Al-Zahery N, Battaglia V, et al. Origin, diffusion, and differentiation of Y-chromosome haplogroups E and J: inferences on the neolithization of Europe and later migratory events in the Mediterranean area. Am J Hum Genet 2004 May;74(5): (204) Underhill PA, Passarino G, Lin AA, Shen P, Mirazon LM, Foley RA, et al. The phylogeography of Y chromosome binary haplotypes and the origins of modern human populations. Ann Hum Genet 2001 Jan;65(Pt 1): (205) Carvalho-Silva DR, Santos FR, Rocha J, Pena SD. The phylogeography of Brazilian Y-chromosome lineages. Am J Hum Genet 2001 Jan;68(1): (206) Carvalho-Silva DR, Tarazona-Santos E, Rocha J, Pena SD, Santos FR. Y chromosome diversity in Brazilians: switching perspectives from slow to fast evolving markers. Genetica 2006 Jan;126(1-2): (207) Cockerham CC, Weir BS. Covariances of relatives stemming from a population undergoing mixed self and random mating. Biometrics 1984 Mar;40(1): (208) Excoffier L, Smouse PE, Quattro JM. Analysis of molecular variance inferred from metric distances among DNA haplotypes: application to human mitochondrial DNA restriction data. Genetics 1992 Jun;131(2): (209) Excoffier L, Heckel G. Computer programs for population genetics data analysis: a survival guide. Nat Rev Genet 2006 Oct;7(10): (210) Dereure J, Duong TH, Lavabre-Bertrand T, Cartron G, Bastides F, Richard-Lenoble D, et al. Visceral leishmaniasis. Persistence of parasites in lymph nodes after clinical cure. J Infect 2003 Jul;47(1): (211) Schubach A, Haddad F, Oliveira-Neto MP, Degrave W, Pirmez C, Grimaldi G, Jr., et al. Detection of Leishmania DNA by polymerase chain reaction in scars of treated human patients. J Infect Dis 1998 Sep;178(3): (212) Schubach A, Marzochi MC, Cuzzi-Maya T, Oliveira AV, Araujo ML, Oliveira AL, et al. Cutaneous scars in American tegumentary leishmaniasis patients: a site of Leishmania (Viannia) braziliensis 86
91 persistence and viability eleven years after antimonial therapy and clinical cure. Am J Trop Med Hyg 1998 Jun;58(6): (213) Bucheton B, Abel L, El-Safi S, Kheir MM, Pavek S, Lemainque A, et al. A major susceptibility locus on chromosome 22q12 plays a critical role in the control of kala-azar. Am J Hum Genet 2003 Nov;73(5): (214) Miller EN, Fadl M, Mohamed HS, Elzein A, Jamieson SE, Cordell HJ, et al. Y chromosome lineage- and village-specific genes on chromosomes 1p22 and 6q27 control visceral leishmaniasis in Sudan. PLoS Genet 2007 May 11;3(5):e71. (215) Khalil EA, Zijlstra EE, Kager PA, El Hassan AM. Epidemiology and clinical manifestations of Leishmania donovani infection in two villages in an endemic area in eastern Sudan. Trop Med Int Health 2002 Jan;7(1): (216) Jamieson SE, Miller EN, Peacock CS, Fakiola M, Wilson ME, Bales- Holst A, et al. Genome-wide scan for visceral leishmaniasis susceptibility genes in Brazil. Genes Immun 2007 Jan;8(1):

92 8. ANEXO 88

93 89

94 90
95 91
96 92

97 93
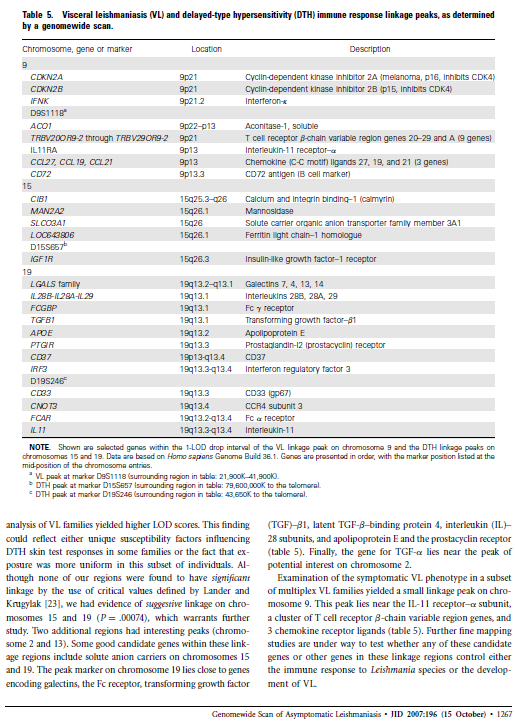
98 94

99 95

100 96
Portuguese Summary. Resumo
 Portuguese Summary Resumo 176 Resumo Cerca de 1 em 100 indivíduos não podem comer pão, macarrão ou biscoitos, pois eles têm uma condição chamada de doença celíaca (DC). DC é causada por uma das intolerâncias
Portuguese Summary Resumo 176 Resumo Cerca de 1 em 100 indivíduos não podem comer pão, macarrão ou biscoitos, pois eles têm uma condição chamada de doença celíaca (DC). DC é causada por uma das intolerâncias
Diagnóstico Microbiológico
 Diagnóstico Microbiológico Identificação e Tipagem Bacteriana Prof. Vânia Lúcia Diagnóstico clínico Sinais (mensuráveis) e sintomas (subjetivos) Origem Etiologia Natureza Diagnóstico laboratorial Identificação
Diagnóstico Microbiológico Identificação e Tipagem Bacteriana Prof. Vânia Lúcia Diagnóstico clínico Sinais (mensuráveis) e sintomas (subjetivos) Origem Etiologia Natureza Diagnóstico laboratorial Identificação
Pesquisa inédita avalia conhecimento da população sobre a tuberculose
 Pesquisa inédita avalia conhecimento da população sobre a tuberculose Uma pesquisa quantitativa de opinião pública realizada pelo Núcleo de Pesquisas da Universidade Federal Fluminense (DataUFF) demonstra
Pesquisa inédita avalia conhecimento da população sobre a tuberculose Uma pesquisa quantitativa de opinião pública realizada pelo Núcleo de Pesquisas da Universidade Federal Fluminense (DataUFF) demonstra
BIOLOGIA - 3 o ANO MÓDULO 28 REINO PROTISTA
 BIOLOGIA - 3 o ANO MÓDULO 28 REINO PROTISTA Corpúsculo basal Membrana ondulante Citoplasma núcleo flagelo Reprodução assexuada em flagelados Parasita Leishmania brasiliensis Transmissor homem Phlebotomus
BIOLOGIA - 3 o ANO MÓDULO 28 REINO PROTISTA Corpúsculo basal Membrana ondulante Citoplasma núcleo flagelo Reprodução assexuada em flagelados Parasita Leishmania brasiliensis Transmissor homem Phlebotomus
LISTA DE VERIFICAÇAO DO SISTEMA DE GESTAO DA QUALIDADE
 Questionamento a alta direção: 1. Quais os objetivos e metas da organização? 2. quais os principais Produtos e/ou serviços da organização? 3. Qual o escopo da certificação? 4. qual é a Visão e Missão?
Questionamento a alta direção: 1. Quais os objetivos e metas da organização? 2. quais os principais Produtos e/ou serviços da organização? 3. Qual o escopo da certificação? 4. qual é a Visão e Missão?
GENÉTICA DE POPULAÇÕES:
 Genética Animal Fatores Evolutivos 1 GENÉTICA DE POPULAÇÕES: A genética de populações lida com populações naturais. Estas consistem em todos os indivíduos que, ao se reproduzir uns com os outros, compartilham
Genética Animal Fatores Evolutivos 1 GENÉTICA DE POPULAÇÕES: A genética de populações lida com populações naturais. Estas consistem em todos os indivíduos que, ao se reproduzir uns com os outros, compartilham
Copyright Proibida Reprodução. Prof. Éder Clementino dos Santos
 NOÇÕES DE OHSAS 18001:2007 CONCEITOS ELEMENTARES SISTEMA DE GESTÃO DE SSO OHSAS 18001:2007? FERRAMENTA ELEMENTAR CICLO DE PDCA (OHSAS 18001:2007) 4.6 ANÁLISE CRÍTICA 4.3 PLANEJAMENTO A P C D 4.5 VERIFICAÇÃO
NOÇÕES DE OHSAS 18001:2007 CONCEITOS ELEMENTARES SISTEMA DE GESTÃO DE SSO OHSAS 18001:2007? FERRAMENTA ELEMENTAR CICLO DE PDCA (OHSAS 18001:2007) 4.6 ANÁLISE CRÍTICA 4.3 PLANEJAMENTO A P C D 4.5 VERIFICAÇÃO
Entre 1998 e 2001, a freqüência escolar aumentou bastante no Brasil. Em 1998, 97% das
 INFORME-SE BNDES ÁREA PARA ASSUNTOS FISCAIS E DE EMPREGO AFE Nº 48 NOVEMBRO DE 2002 EDUCAÇÃO Desempenho educacional no Brasil: O que nos diz a PNAD-2001 Entre 1998 e 2001, a freqüência escolar aumentou
INFORME-SE BNDES ÁREA PARA ASSUNTOS FISCAIS E DE EMPREGO AFE Nº 48 NOVEMBRO DE 2002 EDUCAÇÃO Desempenho educacional no Brasil: O que nos diz a PNAD-2001 Entre 1998 e 2001, a freqüência escolar aumentou
BIOFÍSICA DAS RADIAÇÕES IONIZANTES
 BIOFÍSICA DAS RADIAÇÕES IONIZANTES DANOS RADIOINDUZIDOS NA MOLÉCULA DE DNA Por ser responsável pela codificação da estrutura molecular de todas as enzimas da células, o DNA passa a ser a molécula chave
BIOFÍSICA DAS RADIAÇÕES IONIZANTES DANOS RADIOINDUZIDOS NA MOLÉCULA DE DNA Por ser responsável pela codificação da estrutura molecular de todas as enzimas da células, o DNA passa a ser a molécula chave
Programa de Certificação de Alimentação Humana, Animal e de Produção de Sementes NÃO OGM
 Programa de Certificação de Alimentação Humana, Animal e de Produção de Sementes NÃO OGM* (*organismos geneticamente modificados) 2ª Edição - Maio 2015 2 ÍNDICE 1. INTRODUÇÃO... 3 2. DEFINIÇÃO DO ESCOPO...
Programa de Certificação de Alimentação Humana, Animal e de Produção de Sementes NÃO OGM* (*organismos geneticamente modificados) 2ª Edição - Maio 2015 2 ÍNDICE 1. INTRODUÇÃO... 3 2. DEFINIÇÃO DO ESCOPO...
www.printo.it/pediatric-rheumatology/br/intro
 www.printo.it/pediatric-rheumatology/br/intro Artrite de lyme Versão de 2016 1. O QUE É ARTRITE DE LYME 1.1 O que é? A artrite de Lyme é uma das doenças causadas pela bactéria Borrelia burgdorferi (borreliose
www.printo.it/pediatric-rheumatology/br/intro Artrite de lyme Versão de 2016 1. O QUE É ARTRITE DE LYME 1.1 O que é? A artrite de Lyme é uma das doenças causadas pela bactéria Borrelia burgdorferi (borreliose
Conhecendo o vírus v. Vírus da Imunodeficiência Humana VIH
 Conhecendo o vírus v da Sida Vírus da Imunodeficiência Humana VIH Conhecendo o Vírus da Sida O vírus entra na corrente sanguínea; Determina a posição exacta e reconhece os linfócitos T helper, fixando-se
Conhecendo o vírus v da Sida Vírus da Imunodeficiência Humana VIH Conhecendo o Vírus da Sida O vírus entra na corrente sanguínea; Determina a posição exacta e reconhece os linfócitos T helper, fixando-se
BRASCAN RESIDENTIAL PROPERTIES S.A. PLANO DE OPÇÃO DE COMPRA DE AÇÕES
 BRASCAN RESIDENTIAL PROPERTIES S.A. PLANO DE OPÇÃO DE COMPRA DE AÇÕES SUMÁRIO PLANO DE OPÇÃO PARA AÇÃO... 3 SEÇÃO 1. DISPOSIÇÃO GERAL... 3 1.1 Propósito... 3 1.2 Administração... 3 1.3 Interpretação...
BRASCAN RESIDENTIAL PROPERTIES S.A. PLANO DE OPÇÃO DE COMPRA DE AÇÕES SUMÁRIO PLANO DE OPÇÃO PARA AÇÃO... 3 SEÇÃO 1. DISPOSIÇÃO GERAL... 3 1.1 Propósito... 3 1.2 Administração... 3 1.3 Interpretação...
A Meta-Analytic Review of Psychosocial Interventions for Substance Use Disorders
 A Meta-Analytic Review of Psychosocial Interventions for Substance Use Disorders REVISÃO META-ANALÍTICA DO USO DE INTERVENÇÕES PSICOSSOCIAIS NO TRATAMENTO DE DEPENDÊNCIA QUÍMICA Publicado: Am J Psychiattry
A Meta-Analytic Review of Psychosocial Interventions for Substance Use Disorders REVISÃO META-ANALÍTICA DO USO DE INTERVENÇÕES PSICOSSOCIAIS NO TRATAMENTO DE DEPENDÊNCIA QUÍMICA Publicado: Am J Psychiattry
DIRETRIZES E PARÂMETROS DE AVALIAÇÃO DE PROPOSTAS DE CURSOS NOVOS DE MESTRADO PROFISSIONAL
 DIRETRIZES E PARÂMETROS DE AVALIAÇÃO DE PROPOSTAS DE CURSOS NOVOS DE MESTRADO PROFISSIONAL I) Apresentação Este documento descreve as diretrizes e parâmetros de avaliação de mestrado profissional em Administração,
DIRETRIZES E PARÂMETROS DE AVALIAÇÃO DE PROPOSTAS DE CURSOS NOVOS DE MESTRADO PROFISSIONAL I) Apresentação Este documento descreve as diretrizes e parâmetros de avaliação de mestrado profissional em Administração,
Uso de Substâncias Psicoativas
 Uso de Substâncias Psicoativas X Direção Veicular ALOISIO ANDRADE Psiquiatra e Homeopata XI Jornada Mineira de Medicina de Tráfego Belo Horizonte - MG 18 e 19/07/2014 I-Dados Estatísticos - O Brasil ocupa
Uso de Substâncias Psicoativas X Direção Veicular ALOISIO ANDRADE Psiquiatra e Homeopata XI Jornada Mineira de Medicina de Tráfego Belo Horizonte - MG 18 e 19/07/2014 I-Dados Estatísticos - O Brasil ocupa
ECONTEXTO. Auditoria Ambiental e de Regularidade
 Auditoria Ambiental e de Regularidade Organização Internacional das Entidades Fiscalizadoras Superiores - INTOSAI Grupo de Trabalho sobre Auditoria Ambiental - WGEA ECONTEXTO Este artigo é um resumo do
Auditoria Ambiental e de Regularidade Organização Internacional das Entidades Fiscalizadoras Superiores - INTOSAI Grupo de Trabalho sobre Auditoria Ambiental - WGEA ECONTEXTO Este artigo é um resumo do
PERFIL DOS RADIOLOGISTAS NO BRASIL: análise dos dados INTRODUÇÃO
 1 PERFIL DOS RADIOLOGISTAS NO BRASIL: análise dos dados INTRODUÇÃO O Brasil conta hoje, com 254.886 médicos em atividade profissional (CFM, 2003). O contingente de radiologistas é da ordem de 5388, o que
1 PERFIL DOS RADIOLOGISTAS NO BRASIL: análise dos dados INTRODUÇÃO O Brasil conta hoje, com 254.886 médicos em atividade profissional (CFM, 2003). O contingente de radiologistas é da ordem de 5388, o que
Q U E S T Ã O 4 0 Q U E S T Ã O 4 1 PROVA DE BIOLOGIA II
 23 PROVA DE BIOLOGIA II Q U E S T Ã O 4 0 Cientistas americanos conseguiram modificar geneticamente bichos-da-seda para fazê-los produzir teia de aranha, um material conhecido por sua resistência e elasticidade,
23 PROVA DE BIOLOGIA II Q U E S T Ã O 4 0 Cientistas americanos conseguiram modificar geneticamente bichos-da-seda para fazê-los produzir teia de aranha, um material conhecido por sua resistência e elasticidade,
Auto-imunidade Doenças auto-imunes. Sandra Bertelli Ribeiro sandrabertelliribeiro@hotmail.com Doutoranda Lab. de Imunologia
 Auto-imunidade Doenças auto-imunes Sandra Bertelli Ribeiro sandrabertelliribeiro@hotmail.com Doutoranda Lab. de Imunologia Célula tronco-hematopoiética Pluripotente. - Progenitor linfóide comum - Progenitor
Auto-imunidade Doenças auto-imunes Sandra Bertelli Ribeiro sandrabertelliribeiro@hotmail.com Doutoranda Lab. de Imunologia Célula tronco-hematopoiética Pluripotente. - Progenitor linfóide comum - Progenitor
Introdução à Filogenética para Professores de Biologia
 Introdução à Filogenética para Professores de Biologia A filogenética ajuda a organizar a grande biodiversidade de forma a evidenciar a Evolução. Vamos entender como isso funciona? www.botanicaonline.com.br
Introdução à Filogenética para Professores de Biologia A filogenética ajuda a organizar a grande biodiversidade de forma a evidenciar a Evolução. Vamos entender como isso funciona? www.botanicaonline.com.br
Segurança e Saúde dos Trabalhadores
 Segurança e Saúde dos Trabalhadores [1]CONVENÇÃO N. 155 I Aprovada na 67ª reunião da Conferência Internacional do Trabalho (Genebra 1981), entrou em vigor no plano internacional em 11.8.83. II Dados referentes
Segurança e Saúde dos Trabalhadores [1]CONVENÇÃO N. 155 I Aprovada na 67ª reunião da Conferência Internacional do Trabalho (Genebra 1981), entrou em vigor no plano internacional em 11.8.83. II Dados referentes
Recomendada. A coleção apresenta eficiência e adequação. Ciências adequados a cada faixa etária, além de
 Recomendada Por quê? A coleção apresenta eficiência e adequação metodológica, com os principais temas relacionados a Ciências adequados a cada faixa etária, além de conceitos em geral corretos. Constitui
Recomendada Por quê? A coleção apresenta eficiência e adequação metodológica, com os principais temas relacionados a Ciências adequados a cada faixa etária, além de conceitos em geral corretos. Constitui
PED ABC Novembro 2015
 PESQUISA DE EMPREGO E DESEMPREGO NA REGIÃO DO ABC 1 Novembro 2015 OS NEGROS NO MERCADO DE TRABALHO DA REGIÃO DO ABC Diferenciais de inserção de negros e não negros no mercado de trabalho em 2013-2014 Dia
PESQUISA DE EMPREGO E DESEMPREGO NA REGIÃO DO ABC 1 Novembro 2015 OS NEGROS NO MERCADO DE TRABALHO DA REGIÃO DO ABC Diferenciais de inserção de negros e não negros no mercado de trabalho em 2013-2014 Dia
ANEXO I TRANSPLANTE DE CÉLULAS-TRONCO HEMATOPOÉTICAS TCTH REGULAMENTO TÉCNICO
 ANEXO I TRANSPLANTE DE CÉLULAS-TRONCO HEMATOPOÉTICAS TCTH REGULAMENTO TÉCNICO I - DEFINIÇÕES 1. TCTH de Medula Óssea - Substituição de células-tronco hematopoéticas a partir de células-tronco hematopoéticas
ANEXO I TRANSPLANTE DE CÉLULAS-TRONCO HEMATOPOÉTICAS TCTH REGULAMENTO TÉCNICO I - DEFINIÇÕES 1. TCTH de Medula Óssea - Substituição de células-tronco hematopoéticas a partir de células-tronco hematopoéticas
Guia de utilização da notação BPMN
 1 Guia de utilização da notação BPMN Agosto 2011 2 Sumário de Informações do Documento Documento: Guia_de_utilização_da_notação_BPMN.odt Número de páginas: 31 Versão Data Mudanças Autor 1.0 15/09/11 Criação
1 Guia de utilização da notação BPMN Agosto 2011 2 Sumário de Informações do Documento Documento: Guia_de_utilização_da_notação_BPMN.odt Número de páginas: 31 Versão Data Mudanças Autor 1.0 15/09/11 Criação
Unidade 9: Diálogos deliberativos
 Unidade 9: Diálogos deliberativos Como podemos utilizar as sínteses de evidências? Informar os grupos de interesse Divulgação da síntese de políticas Informações adaptadas derivadas da síntese Meios de
Unidade 9: Diálogos deliberativos Como podemos utilizar as sínteses de evidências? Informar os grupos de interesse Divulgação da síntese de políticas Informações adaptadas derivadas da síntese Meios de
HEMORIO INSTITUTO ESTADUAL DE HEMATOLOGIA ARTHUR DE SIQUEIRA CAVALCANTI
 MANUAL DO PACIENTE - LEUCEMIA LINFÓIDE AGUDA EDIÇÃO REVISADA 02/2009 HEMORIO INSTITUTO ESTADUAL DE HEMATOLOGIA ARTHUR DE SIQUEIRA CAVALCANTI Este manual tem como objetivo fornecer informações aos pacientes
MANUAL DO PACIENTE - LEUCEMIA LINFÓIDE AGUDA EDIÇÃO REVISADA 02/2009 HEMORIO INSTITUTO ESTADUAL DE HEMATOLOGIA ARTHUR DE SIQUEIRA CAVALCANTI Este manual tem como objetivo fornecer informações aos pacientes
www.fundep.br/programacaptar, juntamente com este regulamento.
 PROGRAMA DE CAPTAÇÃO DE RECURSOS FUNDEP REGULAMENTO PARA CADASTRAMENTO DE PROJETOS UFMG A Fundep//Gerência de Articulação de Parcerias convida a comunidade acadêmica da UFMG a cadastrar propostas de acordo
PROGRAMA DE CAPTAÇÃO DE RECURSOS FUNDEP REGULAMENTO PARA CADASTRAMENTO DE PROJETOS UFMG A Fundep//Gerência de Articulação de Parcerias convida a comunidade acadêmica da UFMG a cadastrar propostas de acordo
Leishmaniose visceral: vacina europeia traz fórmula inovadora
 Leishmaniose visceral: vacina europeia traz fórmula inovadora Em entrevista à Sociedade Brasileira de Medicina Tropical (SBMT), Dr. Javier Moreno fala sobre a vacina contra leishmanioses (http://www.ideal.es/jaen/20140212/mas-actualidad/ciencia/europaimpulsa-vacuna-contra-201402121152.html)
Leishmaniose visceral: vacina europeia traz fórmula inovadora Em entrevista à Sociedade Brasileira de Medicina Tropical (SBMT), Dr. Javier Moreno fala sobre a vacina contra leishmanioses (http://www.ideal.es/jaen/20140212/mas-actualidad/ciencia/europaimpulsa-vacuna-contra-201402121152.html)
VÍRUS (complementar o estudo com as páginas 211-213 do livro texto)
") COLÉGIO E CURSO INTELECTUS APOSTILA NOME: MAT.: Biologia I PROFº: EDUARDO SÉRIE: TURMA: DATA: VÍRUS (complementar o estudo com as páginas 211-213 do livro texto) Os vírus são os únicos organismos acelulares,
COLÉGIO E CURSO INTELECTUS APOSTILA NOME: MAT.: Biologia I PROFº: EDUARDO SÉRIE: TURMA: DATA: VÍRUS (complementar o estudo com as páginas 211-213 do livro texto) Os vírus são os únicos organismos acelulares,
Correntes de Participação e Critérios da Aliança Global Wycliffe [Versão de 9 de maio de 2015]
![Correntes de Participação e Critérios da Aliança Global Wycliffe [Versão de 9 de maio de 2015]](/thumbs/33/16195012.jpg "Correntes de Participação e Critérios da Aliança Global Wycliffe [Versão de 9 de maio de 2015]") Correntes de Participação e Critérios da Aliança Global Wycliffe [Versão de 9 de maio de 2015] Introdução As Organizações da Aliança Global Wycliffe desejam expressar a unidade e a diversidade do Corpo
Correntes de Participação e Critérios da Aliança Global Wycliffe [Versão de 9 de maio de 2015] Introdução As Organizações da Aliança Global Wycliffe desejam expressar a unidade e a diversidade do Corpo
MELHORES PRÁTICAS DA OCDE
 MELHORES PRÁTICAS DA OCDE PARA A TRANSPARÊNCIA ORÇAMENTÁRIA INTRODUÇÃO A relação entre a boa governança e melhores resultados econômicos e sociais é cada vez mais reconhecida. A transparência abertura
MELHORES PRÁTICAS DA OCDE PARA A TRANSPARÊNCIA ORÇAMENTÁRIA INTRODUÇÃO A relação entre a boa governança e melhores resultados econômicos e sociais é cada vez mais reconhecida. A transparência abertura
Origem da variação. Conceitos importantes. Diversidade Genética. Variação genética
 Variação genética Origem da variação Professor Fabrício R Santos fsantos@icb.ufmg.br Departamento de Biologia Geral, UFMG 2012 Variação fenotípica hereditária Variação fenotípica causada pelo ambiente
Variação genética Origem da variação Professor Fabrício R Santos fsantos@icb.ufmg.br Departamento de Biologia Geral, UFMG 2012 Variação fenotípica hereditária Variação fenotípica causada pelo ambiente
O RESULTADO GENÉTICO INCONCLUSIVO PODE DIFICULTAR O DIAGNÓSTICO E A INCLUSÃO DA PESSOA COM DEFICIÊNCIA
 O RESULTADO GENÉTICO INCONCLUSIVO PODE DIFICULTAR O DIAGNÓSTICO E A INCLUSÃO DA PESSOA COM DEFICIÊNCIA Renata Grossi Yuri Lelis Rafael Jessica Cristina Eurich Thais Conceição Silva Wagner José Martins
O RESULTADO GENÉTICO INCONCLUSIVO PODE DIFICULTAR O DIAGNÓSTICO E A INCLUSÃO DA PESSOA COM DEFICIÊNCIA Renata Grossi Yuri Lelis Rafael Jessica Cristina Eurich Thais Conceição Silva Wagner José Martins
ADMINISTRAÇÃO GERAL MOTIVAÇÃO
 ADMINISTRAÇÃO GERAL MOTIVAÇÃO Atualizado em 11/01/2016 MOTIVAÇÃO Estar motivado é visto como uma condição necessária para que um trabalhador entregue um desempenho superior. Naturalmente, como a motivação
ADMINISTRAÇÃO GERAL MOTIVAÇÃO Atualizado em 11/01/2016 MOTIVAÇÃO Estar motivado é visto como uma condição necessária para que um trabalhador entregue um desempenho superior. Naturalmente, como a motivação
PlanetaBio Resolução de Vestibulares UNESP 2010 1ª fase www.planetabio.com
 1- Leia a notícia. Dengue tipo 4 reaparece após 25 anos A dengue é causada por quatro tipos de vírus: DENV-1, DENV-2, DENV-3 e DENV-4. O tipo DENV-4 não era encontrado no país desde 1982, mas exames de
1- Leia a notícia. Dengue tipo 4 reaparece após 25 anos A dengue é causada por quatro tipos de vírus: DENV-1, DENV-2, DENV-3 e DENV-4. O tipo DENV-4 não era encontrado no país desde 1982, mas exames de
Áudio. GUIA DO PROFESSOR Síndrome de Down - Parte I
 Síndrome de Down - Parte I Conteúdos: Tempo: Síndrome de Down 5 minutos Objetivos: Auxiliar o aluno na compreensão do que é síndrome de Down Descrição: Produções Relacionadas: Neste programa de Biologia
Síndrome de Down - Parte I Conteúdos: Tempo: Síndrome de Down 5 minutos Objetivos: Auxiliar o aluno na compreensão do que é síndrome de Down Descrição: Produções Relacionadas: Neste programa de Biologia
Teste seus conhecimentos: Caça-Palavras
 Teste seus conhecimentos: Caça-Palavras Batizada pelos médicos de diabetes mellitus, a doença ocorre quando há um aumento do açúcar no sangue. Dependendo dos motivos desse disparo, pode ser de dois tipos.
Teste seus conhecimentos: Caça-Palavras Batizada pelos médicos de diabetes mellitus, a doença ocorre quando há um aumento do açúcar no sangue. Dependendo dos motivos desse disparo, pode ser de dois tipos.
7.012 Conjunto de Problemas 5
 Nome Seção 7.012 Conjunto de Problemas 5 Pergunta 1 Enquanto estudava um problema de infertilidade, você tentou isolar um gene hipotético de coelho que seria responsável pela prolífica reprodução desses
Nome Seção 7.012 Conjunto de Problemas 5 Pergunta 1 Enquanto estudava um problema de infertilidade, você tentou isolar um gene hipotético de coelho que seria responsável pela prolífica reprodução desses
Estudo PARTNER. Foi convidado a participar neste estudo porque tem uma relação em que é o parceiro VIH positivo.
 Informação ao participante e consentimento informado para o parceiro VIH positivo Estudo PARTNER O estudo PARTNER é um estudo levado a cabo com casais em que: (i) um parceiro é VIH positivo e o outro é
Informação ao participante e consentimento informado para o parceiro VIH positivo Estudo PARTNER O estudo PARTNER é um estudo levado a cabo com casais em que: (i) um parceiro é VIH positivo e o outro é
Trabalho 7 Fila de prioridade usando heap para simulação de atendimento
 Trabalho 7 Fila de prioridade usando heap para simulação de atendimento Data: 21/10/2013 até meia-noite Dúvidas até: 09/10/2013 Faq disponível em: http://www2.icmc.usp.br/~mello/trabalho07.html A estrutura
Trabalho 7 Fila de prioridade usando heap para simulação de atendimento Data: 21/10/2013 até meia-noite Dúvidas até: 09/10/2013 Faq disponível em: http://www2.icmc.usp.br/~mello/trabalho07.html A estrutura
Categorias Temas Significados Propostos
 91 5. Conclusão O objetivo do presente trabalho foi descrever a essência do significado da experiência consultiva para profissionais de TI que prestam de serviços de consultoria na área de TI. Para atingir
91 5. Conclusão O objetivo do presente trabalho foi descrever a essência do significado da experiência consultiva para profissionais de TI que prestam de serviços de consultoria na área de TI. Para atingir
ipea políticas sociais acompanhamento e análise 7 ago. 2003 117 GASTOS SOCIAIS: FOCALIZAR VERSUS UNIVERSALIZAR José Márcio Camargo*
 GASTOS SOCIAIS: FOCALIZAR VERSUS UNIVERSALIZAR José Márcio Camargo* Como deve ser estruturada a política social de um país? A resposta a essa pergunta independe do grau de desenvolvimento do país, da porcentagem
GASTOS SOCIAIS: FOCALIZAR VERSUS UNIVERSALIZAR José Márcio Camargo* Como deve ser estruturada a política social de um país? A resposta a essa pergunta independe do grau de desenvolvimento do país, da porcentagem
Estado da tecnologia avançada na gestão dos recursos genéticos animais
 PARTE 4 Estado da tecnologia avançada na gestão dos recursos genéticos animais A caracterização de raças e ambientes de produção precisa ser melhorada para fomentar políticas de decisão na gestão dos recursos
PARTE 4 Estado da tecnologia avançada na gestão dos recursos genéticos animais A caracterização de raças e ambientes de produção precisa ser melhorada para fomentar políticas de decisão na gestão dos recursos
Inovação aberta na indústria de software: Avaliação do perfil de inovação de empresas
 : Avaliação do perfil de inovação de empresas Prof. Paulo Henrique S. Bermejo, Dr. Prof. André Luiz Zambalde, Dr. Adriano Olímpio Tonelli, MSc. Pamela A. Santos Priscila Rosa LabGTI Laboratório de Governança
: Avaliação do perfil de inovação de empresas Prof. Paulo Henrique S. Bermejo, Dr. Prof. André Luiz Zambalde, Dr. Adriano Olímpio Tonelli, MSc. Pamela A. Santos Priscila Rosa LabGTI Laboratório de Governança
Ferrarezi News. Setembro/2015. News. Ferrarezi. Onda de virose? Tudo é Virose? Programa - PRO Mamãe & Bebê. Depressão
 Setembro/2015 3 Onda de virose? 6 Tudo é Virose? 10 Programa - PRO Mamãe & Bebê 11 Depressão Setembro/2015 Onda de virose? O virologista Celso Granato esclarece Ouço muita gente falar em virose. Procurei
Setembro/2015 3 Onda de virose? 6 Tudo é Virose? 10 Programa - PRO Mamãe & Bebê 11 Depressão Setembro/2015 Onda de virose? O virologista Celso Granato esclarece Ouço muita gente falar em virose. Procurei
Ancestralidade Materna polimorfismos matrilínea DNA Mitocondrial (mtdna).
.") Ancestralidade Materna A atual população dos países latino-americanos foi gerada por um complexo processo de mistura genética entre ameríndios, europeus e africanos. As porcentagens relativas destas três
Ancestralidade Materna A atual população dos países latino-americanos foi gerada por um complexo processo de mistura genética entre ameríndios, europeus e africanos. As porcentagens relativas destas três
ÍNDICE DE QUALIDADE DE VIDA DO TRABALHADOR DA INDÚSTRIA - SESI/SC
 ÍNDICE DE QUALIDADE DE VIDA DO TRABALHADOR DA INDÚSTRIA - SESI/SC SUMÁRIO EXECUTIVO 2015 Apresentação Integrando a agenda mundial para a promoção da saúde e produtividade, o SESI Santa Catarina realizou
ÍNDICE DE QUALIDADE DE VIDA DO TRABALHADOR DA INDÚSTRIA - SESI/SC SUMÁRIO EXECUTIVO 2015 Apresentação Integrando a agenda mundial para a promoção da saúde e produtividade, o SESI Santa Catarina realizou
Política monetária e senhoriagem: depósitos compulsórios na economia brasileira recente
 Política monetária e senhoriagem: depósitos compulsórios na economia brasileira recente Roberto Meurer * RESUMO - Neste artigo se analisa a utilização dos depósitos compulsórios sobre depósitos à vista
Política monetária e senhoriagem: depósitos compulsórios na economia brasileira recente Roberto Meurer * RESUMO - Neste artigo se analisa a utilização dos depósitos compulsórios sobre depósitos à vista
TÍTULO: DETERMINAÇÃO DO INTERVALO DE REFERÊNCIA DOS PRINCIPAIS PARÂMETROS DO HEMOGRAMA PARA HOMENS ADULTOS ATENDIDOS NO LAC-CCF-UNAERP
 TÍTULO: DETERMINAÇÃO DO INTERVALO DE REFERÊNCIA DOS PRINCIPAIS PARÂMETROS DO HEMOGRAMA PARA HOMENS ADULTOS ATENDIDOS NO LAC-CCF-UNAERP CATEGORIA: EM ANDAMENTO ÁREA: CIÊNCIAS BIOLÓGICAS E SAÚDE SUBÁREA:
TÍTULO: DETERMINAÇÃO DO INTERVALO DE REFERÊNCIA DOS PRINCIPAIS PARÂMETROS DO HEMOGRAMA PARA HOMENS ADULTOS ATENDIDOS NO LAC-CCF-UNAERP CATEGORIA: EM ANDAMENTO ÁREA: CIÊNCIAS BIOLÓGICAS E SAÚDE SUBÁREA:
A presente seção apresenta e especifica as hipótese que se buscou testar com o experimento. A seção 5 vai detalhar o desenho do experimento.
 4 Plano de Análise O desenho do experimento realizado foi elaborado de forma a identificar o quão relevantes para a explicação do fenômeno de overbidding são os fatores mencionados na literatura em questão
4 Plano de Análise O desenho do experimento realizado foi elaborado de forma a identificar o quão relevantes para a explicação do fenômeno de overbidding são os fatores mencionados na literatura em questão
ANÁLISE DOS RESULTADOS DOS PROGRAMAS DE APOIO ÀS PMEs NO BRASIL Resumo Executivo PARA BAIXAR A AVALIAÇÃO COMPLETA: WWW.IADB.
 ANÁLISE DOS RESULTADOS DOS PROGRAMAS DE APOIO ÀS PMEs NO BRASIL Resumo Executivo PARA BAIXAR A AVALIAÇÃO COMPLETA: WWW.IADB.ORG/EVALUATION ANÁLISE DOS RESULTADOS DOS PROGRAMAS DE APOIO ÀS PMEs NO BRASIL
ANÁLISE DOS RESULTADOS DOS PROGRAMAS DE APOIO ÀS PMEs NO BRASIL Resumo Executivo PARA BAIXAR A AVALIAÇÃO COMPLETA: WWW.IADB.ORG/EVALUATION ANÁLISE DOS RESULTADOS DOS PROGRAMAS DE APOIO ÀS PMEs NO BRASIL
COMITÊ DE ÉTICA EM PESQUISA DA ESCOLA DE SAÚDE PÚBLICA DO CEARÁ
 COMITÊ DE ÉTICA EM PESQUISA DA ESCOLA DE SAÚDE PÚBLICA DO CEARÁ 1. ORIENTAÇÕES PARA ELABORAÇÃO DO TERMO DE CONSENTIMENTO LIVRE E ESCLARECIDO (TCLE) A importância do TCLE. A Resolução CNS 196/96 afirma
COMITÊ DE ÉTICA EM PESQUISA DA ESCOLA DE SAÚDE PÚBLICA DO CEARÁ 1. ORIENTAÇÕES PARA ELABORAÇÃO DO TERMO DE CONSENTIMENTO LIVRE E ESCLARECIDO (TCLE) A importância do TCLE. A Resolução CNS 196/96 afirma
Modos de vida no município de Paraty - Ponta Negra
 Modos de vida no município de Paraty - Ponta Negra Resultados gerais Dezembro 2010 Projeto Community-based resource management and food security in coastal Brazil (Universidade Estadual de Campinas/UNICAMP)
Modos de vida no município de Paraty - Ponta Negra Resultados gerais Dezembro 2010 Projeto Community-based resource management and food security in coastal Brazil (Universidade Estadual de Campinas/UNICAMP)
Redação do Site Inovação Tecnológica - 28/08/2009. Humanos aprimorados versus humanos comuns
 VOCÊ ESTÁ PREPARADO PARA CONVIVER COM OS HUMANOS APRIMORADOS? http://www.inovacaotecnologica.com.br/noticias/noticia.php?artigo=voce-esta-preparado-conviver-humanosaprimorados&id=010850090828 Redação do
VOCÊ ESTÁ PREPARADO PARA CONVIVER COM OS HUMANOS APRIMORADOS? http://www.inovacaotecnologica.com.br/noticias/noticia.php?artigo=voce-esta-preparado-conviver-humanosaprimorados&id=010850090828 Redação do
PREVISÃO DE DEMANDA - O QUE PREVISÃO DE DEMANDA - TIPOS E TÉCNICAS DE PREVISÃO DE DEMANDA - MÉTODOS DE PREVISÃO - EXERCÍCIOS
 CONTEÚDO DO CURSO DE PREVISÃO DE DEMANDA PROMOVIDO PELA www.administrabrasil.com.br - O QUE PREVISÃO DE DEMANDA - TIPOS E TÉCNICAS DE PREVISÃO DE DEMANDA - MÉTODOS DE PREVISÃO - EXERCÍCIOS - HORIZONTE
CONTEÚDO DO CURSO DE PREVISÃO DE DEMANDA PROMOVIDO PELA www.administrabrasil.com.br - O QUE PREVISÃO DE DEMANDA - TIPOS E TÉCNICAS DE PREVISÃO DE DEMANDA - MÉTODOS DE PREVISÃO - EXERCÍCIOS - HORIZONTE
Aula IV Protozoários Zoopatogênicos
 ZOOLOGIA E PARASITOLOGIA Aula IV Protozoários Zoopatogênicos Professora: Luciana Alves de Sousa Doenças causadas por Protozoários Doença de Chagas Leishmaniose Malária Giardíase Amebíase Toxoplasmose Doença
ZOOLOGIA E PARASITOLOGIA Aula IV Protozoários Zoopatogênicos Professora: Luciana Alves de Sousa Doenças causadas por Protozoários Doença de Chagas Leishmaniose Malária Giardíase Amebíase Toxoplasmose Doença
O DNA é formado por pedaços capazes de serem convertidos em algumas características. Esses pedaços são
 Atividade extra Fascículo 2 Biologia Unidade 4 Questão 1 O DNA é formado por pedaços capazes de serem convertidos em algumas características. Esses pedaços são chamados de genes. Assinale abaixo quais
Atividade extra Fascículo 2 Biologia Unidade 4 Questão 1 O DNA é formado por pedaços capazes de serem convertidos em algumas características. Esses pedaços são chamados de genes. Assinale abaixo quais
RESUMOS DE PROJETOS... 124 ARTIGOS COMPLETOS(RESUMOS)... 128
... 128") 123 RESUMOS DE PROJETOS... 124 ARTIGOS COMPLETOS(RESUMOS)... 128 RESUMOS DE PROJETOS 124 A GENÉTICA E NEUROFISIOLOGIA DO AUTISMO... 125 PAPEL DO POLIMORFISMO IL17A (RS7747909) NA TUBERCULOSE.... 126 PAPEL
123 RESUMOS DE PROJETOS... 124 ARTIGOS COMPLETOS(RESUMOS)... 128 RESUMOS DE PROJETOS 124 A GENÉTICA E NEUROFISIOLOGIA DO AUTISMO... 125 PAPEL DO POLIMORFISMO IL17A (RS7747909) NA TUBERCULOSE.... 126 PAPEL
PROPRIEDADES E VISÃO GERAL DAS RESPOSTAS IMUNES. FARMÁCIA PROFa SIMONE PETRI AULA - 1
 PROPRIEDADES E VISÃO GERAL DAS RESPOSTAS IMUNES FARMÁCIA PROFa SIMONE PETRI AULA - 1 INTRODUÇÃO A função fisiológica do sistema imune é a defesa contra micro-organismos infecciosos. Entretanto, mesmo
PROPRIEDADES E VISÃO GERAL DAS RESPOSTAS IMUNES FARMÁCIA PROFa SIMONE PETRI AULA - 1 INTRODUÇÃO A função fisiológica do sistema imune é a defesa contra micro-organismos infecciosos. Entretanto, mesmo
BASES MACROMOLECULARES DA CONSTITUIÇÃO CELULAR
 BASES MACROMOLECULARES DA CONSTITUIÇÃO CELULAR As moléculas que constituem as células são formadas pelos mesmos átomos encontrados nos seres inanimados. Todavia, na origem e evolução das células, alguns
BASES MACROMOLECULARES DA CONSTITUIÇÃO CELULAR As moléculas que constituem as células são formadas pelos mesmos átomos encontrados nos seres inanimados. Todavia, na origem e evolução das células, alguns
A SEGUIR ALGUMAS DICAS PARA O DESENVOLVIMENTO DE UM PROJETO CIENTÍFICO
 A SEGUIR ALGUMAS DICAS PARA O DESENVOLVIMENTO DE UM PROJETO CIENTÍFICO DESENVOLVENDO UM PROJETO 1. Pense em um tema de seu interesse ou um problema que você gostaria de resolver. 2. Obtenha um caderno
A SEGUIR ALGUMAS DICAS PARA O DESENVOLVIMENTO DE UM PROJETO CIENTÍFICO DESENVOLVENDO UM PROJETO 1. Pense em um tema de seu interesse ou um problema que você gostaria de resolver. 2. Obtenha um caderno
Ensaio de impacto. Os veículos brasileiros têm, em geral, suspensão
 A UU L AL A Ensaio de impacto Os veículos brasileiros têm, em geral, suspensão mais reforçada do que a dos similares europeus. Não é à toa. As condições de nossas estradas e ruas requerem esse reforço,
A UU L AL A Ensaio de impacto Os veículos brasileiros têm, em geral, suspensão mais reforçada do que a dos similares europeus. Não é à toa. As condições de nossas estradas e ruas requerem esse reforço,
a) A diversidade de anticorpos é derivada da recombinação das regiões, e.
 A diversidade de anticorpos é derivada da recombinação das regiões, e.") Questão 1 Preencha as lacunas a) A diversidade de anticorpos é derivada da recombinação das regiões, e. Verdadeiro ou falso. Se falso, altere a declaração de modo a torná-la verdadeira. b) A exposição
Questão 1 Preencha as lacunas a) A diversidade de anticorpos é derivada da recombinação das regiões, e. Verdadeiro ou falso. Se falso, altere a declaração de modo a torná-la verdadeira. b) A exposição
9 Como o aluno (pré)adolescente vê o livro didático de inglês
adolescente vê o livro didático de inglês") Cap. 9 Como o aluno (pré)adolescente vê o livro didático de inglês 92 9 Como o aluno (pré)adolescente vê o livro didático de inglês Nesta parte do trabalho, analisarei alguns resultados da análise dos
Cap. 9 Como o aluno (pré)adolescente vê o livro didático de inglês 92 9 Como o aluno (pré)adolescente vê o livro didático de inglês Nesta parte do trabalho, analisarei alguns resultados da análise dos
ESTRESSE OCUPACIONAL SAÚDE E SEGURANÇA DO TRABALHO
 ESTRESSE SAÚDE E SEGURANÇA DO TRABALHO Página 1 de 9 1. OBJETIVO... 3 2. ESCOPO... 3 3. DEFINIÇÕES... 4 4. ESTRESSE OCUPACIONAL: CARACTERIZAÇÃO... 4 4.1. Conceitos fundamentais... 4 4.2. Conseqüências
ESTRESSE SAÚDE E SEGURANÇA DO TRABALHO Página 1 de 9 1. OBJETIVO... 3 2. ESCOPO... 3 3. DEFINIÇÕES... 4 4. ESTRESSE OCUPACIONAL: CARACTERIZAÇÃO... 4 4.1. Conceitos fundamentais... 4 4.2. Conseqüências
Planejamento - 7. Planejamento do Gerenciamento do Risco Identificação dos riscos. Mauricio Lyra, PMP
 Planejamento - 7 Planejamento do Gerenciamento do Risco Identificação dos riscos 1 O que é risco? Evento que representa uma ameaça ou uma oportunidade em potencial Plano de gerenciamento do risco Especifica
Planejamento - 7 Planejamento do Gerenciamento do Risco Identificação dos riscos 1 O que é risco? Evento que representa uma ameaça ou uma oportunidade em potencial Plano de gerenciamento do risco Especifica
Gripe pandémica. Factos que deve conhecer acerca da gripe pandémica
 Gripe pandémica Factos que deve conhecer acerca da gripe pandémica De que trata esta brochura? Esta brochura apresenta informações acerca da gripe pandémica. Dá resposta a 10 perguntas importantes acerca
Gripe pandémica Factos que deve conhecer acerca da gripe pandémica De que trata esta brochura? Esta brochura apresenta informações acerca da gripe pandémica. Dá resposta a 10 perguntas importantes acerca
Redução de Homicídios no Brasil
 Ministério da Saúde MS Secretaria de Vigilância em Saúde - SVS Redução de Homicídios no Brasil SUMÁRIO INTRODUÇÃO... 1 METODOLOGIA DE ANÁLISE... 1 RESULTADOS... 2 Homicídios no Brasil... 2 Óbitos por Arma
Ministério da Saúde MS Secretaria de Vigilância em Saúde - SVS Redução de Homicídios no Brasil SUMÁRIO INTRODUÇÃO... 1 METODOLOGIA DE ANÁLISE... 1 RESULTADOS... 2 Homicídios no Brasil... 2 Óbitos por Arma
Gerenciamento de Projetos Modulo VIII Riscos
 Gerenciamento de Projetos Modulo VIII Riscos Prof. Walter Cunha falecomigo@waltercunha.com http://waltercunha.com Bibliografia* Project Management Institute. Conjunto de Conhecimentos em Gerenciamento
Gerenciamento de Projetos Modulo VIII Riscos Prof. Walter Cunha falecomigo@waltercunha.com http://waltercunha.com Bibliografia* Project Management Institute. Conjunto de Conhecimentos em Gerenciamento
CAPÍTULO 25 COERÊNCIA REGULATÓRIA
 CAPÍTULO 25 COERÊNCIA REGULATÓRIA Artigo 25.1: Definições Para efeito deste Capítulo: medida regulatória coberta significa a medida regulatória determinada por cada Parte a ser objeto deste Capítulo nos
CAPÍTULO 25 COERÊNCIA REGULATÓRIA Artigo 25.1: Definições Para efeito deste Capítulo: medida regulatória coberta significa a medida regulatória determinada por cada Parte a ser objeto deste Capítulo nos
5 Instrução e integração
 SEÇÃO 5 Instrução e integração no meio de trabalho Quando um novo funcionário entra para uma organização, é importante que ele receba um bom apoio para entender sua função e a organização. Instrução é
SEÇÃO 5 Instrução e integração no meio de trabalho Quando um novo funcionário entra para uma organização, é importante que ele receba um bom apoio para entender sua função e a organização. Instrução é
TEXTO 7: DELINEAMENTOS PRÉ-EXPERIMENTAIS 1
 1 Laboratório de Psicologia Experimental Departamento de Psicologia UFSJ Disciplina: Método de Pesquisa Quantitativa Professora: Marina Bandeira TEXTO 7: DELINEAMENTOS PRÉ-EXPERIMENTAIS 1 Autores: Selltiz
1 Laboratório de Psicologia Experimental Departamento de Psicologia UFSJ Disciplina: Método de Pesquisa Quantitativa Professora: Marina Bandeira TEXTO 7: DELINEAMENTOS PRÉ-EXPERIMENTAIS 1 Autores: Selltiz
ESTUDO DA PREVALÊNCIA DO CÂNCER BUCAL NO HC DA UNIVERSIDADE FEDERAL DE UBERLÂNDIA, ATRAVÉS DO CID 10
 UNIVERSIDADE FEDERAL DE UBERLÂNDIA 4ª Semana do Servidor e 5ª Semana Acadêmica 2008 UFU 30 anos ESTUDO DA PREVALÊNCIA DO CÂNCER BUCAL NO HC DA UNIVERSIDADE FEDERAL DE UBERLÂNDIA, ATRAVÉS DO CID 10 Leana
UNIVERSIDADE FEDERAL DE UBERLÂNDIA 4ª Semana do Servidor e 5ª Semana Acadêmica 2008 UFU 30 anos ESTUDO DA PREVALÊNCIA DO CÂNCER BUCAL NO HC DA UNIVERSIDADE FEDERAL DE UBERLÂNDIA, ATRAVÉS DO CID 10 Leana
Epidemiologia da Transmissão Vertical do HIV no Brasil
 Epidemiologia da Transmissão Vertical do HIV no Brasil Letícia Legay Vermelho*, Luíza de Paiva Silva* e Antonio José Leal Costa** Introdução A transmissão vertical, também denominada materno-infantil,
Epidemiologia da Transmissão Vertical do HIV no Brasil Letícia Legay Vermelho*, Luíza de Paiva Silva* e Antonio José Leal Costa** Introdução A transmissão vertical, também denominada materno-infantil,
INVESTIGAÇÕES EM TECNOVIGILÂNCIA
 INVESTIGAÇÕES EM TECNOVIGILÂNCIA A principal finalidade da investigação de acidentes e / ou das reações adversas relacionados aos produtos de saúde, em especial no caso da Tecnovigilância, os equipamentos,
INVESTIGAÇÕES EM TECNOVIGILÂNCIA A principal finalidade da investigação de acidentes e / ou das reações adversas relacionados aos produtos de saúde, em especial no caso da Tecnovigilância, os equipamentos,
Evolução da população do Rio Grande do Sul. Maria de Lourdes Teixeira Jardim Fundação de Economia e Estatística. 1 - Introdução
 Evolução da população do Rio Grande do Sul. Maria de Lourdes Teixeira Jardim Fundação de Economia e Estatística Área Temática: Emprego e Mercado de Trabalho, Demografia Econômica. 1 - Introdução Este texto
Evolução da população do Rio Grande do Sul. Maria de Lourdes Teixeira Jardim Fundação de Economia e Estatística Área Temática: Emprego e Mercado de Trabalho, Demografia Econômica. 1 - Introdução Este texto
Perguntas e Respostas sobre o vírus A(H7N9)* Questions and answers about the vírus A(H7N9)
* Questions and answers about the vírus A(H7N9)") Republicação de Artigo * Questions and answers about the vírus A(H7N9) Centers for Disease Control and Prevention Uma nova cepa do vírus da influenza aviária foi descoberta em aves e pessoas na China.
Republicação de Artigo * Questions and answers about the vírus A(H7N9) Centers for Disease Control and Prevention Uma nova cepa do vírus da influenza aviária foi descoberta em aves e pessoas na China.
Aula 4 Estatística Conceitos básicos
 Aula 4 Estatística Conceitos básicos Plano de Aula Amostra e universo Média Variância / desvio-padrão / erro-padrão Intervalo de confiança Teste de hipótese Amostra e Universo A estatística nos ajuda a
Aula 4 Estatística Conceitos básicos Plano de Aula Amostra e universo Média Variância / desvio-padrão / erro-padrão Intervalo de confiança Teste de hipótese Amostra e Universo A estatística nos ajuda a
ECOLOGIA GERAL ECOLOGIA DE POPULAÇÕES (DINÂMICA POPULACIONAL E DISPERSÃO)
") Aula de hoje: ECOLOGIA GERAL ECOLOGIA DE POPULAÇÕES (DINÂMICA POPULACIONAL E DISPERSÃO) Aula 07 Antes de iniciarmos os estudos sobre populações e seus componentes precisamos conhecer e conceituar as estruturas
Aula de hoje: ECOLOGIA GERAL ECOLOGIA DE POPULAÇÕES (DINÂMICA POPULACIONAL E DISPERSÃO) Aula 07 Antes de iniciarmos os estudos sobre populações e seus componentes precisamos conhecer e conceituar as estruturas
Procedimento dos Défices Excessivos (2ª Notificação de 2014)
") Procedimento dos Défices Excessivos 2ª Notificação 2014 30 de setembro de 2014 Procedimento dos Défices Excessivos (2ª Notificação de 2014) Nos termos dos Regulamentos da União Europeia, o INE envia hoje
Procedimento dos Défices Excessivos 2ª Notificação 2014 30 de setembro de 2014 Procedimento dos Défices Excessivos (2ª Notificação de 2014) Nos termos dos Regulamentos da União Europeia, o INE envia hoje
COMENTÁRIO DA PROVA DE BIOLOGIA
 COMENTÁRIO DA PROVA DE BIOLOGIA A prova de Biologia da UFPR apresentou uma boa distribuição de conteúdos ao longo das nove questões. O grau de dificuldade variou entre questões médias e fáceis, o que está
COMENTÁRIO DA PROVA DE BIOLOGIA A prova de Biologia da UFPR apresentou uma boa distribuição de conteúdos ao longo das nove questões. O grau de dificuldade variou entre questões médias e fáceis, o que está
Processos de gerenciamento de projetos em um projeto
 Processos de gerenciamento de projetos em um projeto O gerenciamento de projetos é a aplicação de conhecimentos, habilidades, ferramentas e técnicas às atividades do projeto a fim de cumprir seus requisitos.
Processos de gerenciamento de projetos em um projeto O gerenciamento de projetos é a aplicação de conhecimentos, habilidades, ferramentas e técnicas às atividades do projeto a fim de cumprir seus requisitos.
Primeira e Segunda Lei de Mendel, Polialelia, Sangue e Sexo
 Primeira e Segunda Lei de Mendel, Polialelia, Sangue e Sexo 1. Em uma espécie de planta, a forma dos frutos pode ser alongada, oval ou redonda. Foram realizados quatro tipos de cruzamento entre plantas
Primeira e Segunda Lei de Mendel, Polialelia, Sangue e Sexo 1. Em uma espécie de planta, a forma dos frutos pode ser alongada, oval ou redonda. Foram realizados quatro tipos de cruzamento entre plantas
DESCRIÇÃO DAS PRÁTICAS DE GESTÃO DA INICIATIVA
 DESCRIÇÃO DAS PRÁTICAS DE GESTÃO DA INICIATIVA Como é sabido existe um consenso de que é necessário imprimir qualidade nas ações realizadas pela administração pública. Para alcançar esse objetivo, pressupõe-se
DESCRIÇÃO DAS PRÁTICAS DE GESTÃO DA INICIATIVA Como é sabido existe um consenso de que é necessário imprimir qualidade nas ações realizadas pela administração pública. Para alcançar esse objetivo, pressupõe-se
Programa de Controle da Dengue/SC
 Programa de Controle da Dengue/SC Estratégia operacional de prevenção e controle da dengue para municípios não infestados por Aedes aegypti, infestados por Aedes aegypti sem circulação viral e infestados
Programa de Controle da Dengue/SC Estratégia operacional de prevenção e controle da dengue para municípios não infestados por Aedes aegypti, infestados por Aedes aegypti sem circulação viral e infestados
Avaliação Econômica. Relação entre Desempenho Escolar e os Salários no Brasil
 Avaliação Econômica Relação entre Desempenho Escolar e os Salários no Brasil Objetivo da avaliação: identificar o impacto do desempenho dos brasileiros na Educação Básica em sua renda futura. Dimensões
Avaliação Econômica Relação entre Desempenho Escolar e os Salários no Brasil Objetivo da avaliação: identificar o impacto do desempenho dos brasileiros na Educação Básica em sua renda futura. Dimensões
A Era dos Mosquitos Transgênicos
 A Era dos Mosquitos Transgênicos Depois das plantas geneticamente modificadas, a ciência dá o passo seguinte - e cria um animal transgênico. Seus inventores querem liberá-lo no Brasil. Será que isso é
A Era dos Mosquitos Transgênicos Depois das plantas geneticamente modificadas, a ciência dá o passo seguinte - e cria um animal transgênico. Seus inventores querem liberá-lo no Brasil. Será que isso é
Cotagem de dimensões básicas
 Cotagem de dimensões básicas Introdução Observe as vistas ortográficas a seguir. Com toda certeza, você já sabe interpretar as formas da peça representada neste desenho. E, você já deve ser capaz de imaginar
Cotagem de dimensões básicas Introdução Observe as vistas ortográficas a seguir. Com toda certeza, você já sabe interpretar as formas da peça representada neste desenho. E, você já deve ser capaz de imaginar
Complexo principal de histocompatibilidade
 Complexo principal de histocompatibilidade Todas as espécies possuem um conjunto de genes denominado MHC, cujos produtos são de importância para o reconhecimento intercelular e a discriminação do que é
Complexo principal de histocompatibilidade Todas as espécies possuem um conjunto de genes denominado MHC, cujos produtos são de importância para o reconhecimento intercelular e a discriminação do que é
Boletim Epidemiológico UHE Santo Antônio do Jari
 Editorial Índice - Editorial - Doença Leishmaniose - Gráfico de Notificações - Doença Malária Este é o segundo número do ano de, com veiculação semestral, referente aos meses de janeiro a junho, contendo
Editorial Índice - Editorial - Doença Leishmaniose - Gráfico de Notificações - Doença Malária Este é o segundo número do ano de, com veiculação semestral, referente aos meses de janeiro a junho, contendo
Estrutura para a avaliação de estratégias fiscais para Certificação Empresas B
 Estrutura para a avaliação de estratégias fiscais para Certificação Empresas B Este documento fornece a estrutura que B Lab utiliza para avaliar as estratégias fiscais que atendam aos requisitos da Certificação
Estrutura para a avaliação de estratégias fiscais para Certificação Empresas B Este documento fornece a estrutura que B Lab utiliza para avaliar as estratégias fiscais que atendam aos requisitos da Certificação
b) Ao longo da sucessão ecológica de uma floresta pluvial tropical, restaurada rumo ao clímax, discuta o que ocorre com os seguintes fatores
 Ao longo da sucessão ecológica de uma floresta pluvial tropical, restaurada rumo ao clímax, discuta o que ocorre com os seguintes fatores") Questão 1 Leia o seguinte texto: Com a oportunidade de colocar em prática a nova lei do código florestal brasileiro (lei 12.631/12) e estabelecer estratégias para a recuperação de áreas degradadas, o Ministério
Questão 1 Leia o seguinte texto: Com a oportunidade de colocar em prática a nova lei do código florestal brasileiro (lei 12.631/12) e estabelecer estratégias para a recuperação de áreas degradadas, o Ministério
3 Qualidade de Software
 3 Qualidade de Software Este capítulo tem como objetivo esclarecer conceitos relacionados à qualidade de software; conceitos estes muito importantes para o entendimento do presente trabalho, cujo objetivo
3 Qualidade de Software Este capítulo tem como objetivo esclarecer conceitos relacionados à qualidade de software; conceitos estes muito importantes para o entendimento do presente trabalho, cujo objetivo
3.4 O Princípio da Equipartição de Energia e a Capacidade Calorífica Molar
 3.4 O Princípio da Equipartição de Energia e a Capacidade Calorífica Molar Vimos que as previsões sobre as capacidades caloríficas molares baseadas na teoria cinética estão de acordo com o comportamento
3.4 O Princípio da Equipartição de Energia e a Capacidade Calorífica Molar Vimos que as previsões sobre as capacidades caloríficas molares baseadas na teoria cinética estão de acordo com o comportamento
Juliana Cerqueira de Paiva. Modelos Atômicos Aula 2
 Juliana Cerqueira de Paiva Modelos Atômicos Aula 2 2 Modelo Atômico de Thomson Joseph John Thomson (1856 1940) Por volta de 1897, realizou experimentos estudando descargas elétricas em tubos semelhantes
Juliana Cerqueira de Paiva Modelos Atômicos Aula 2 2 Modelo Atômico de Thomson Joseph John Thomson (1856 1940) Por volta de 1897, realizou experimentos estudando descargas elétricas em tubos semelhantes
Áudio GUIA DO PROFESSOR. Idéias evolucionistas e evolução biológica
 Teoria sintética da evolução Parte II Conteúdos: Tempo: Objetivos: Idéias evolucionistas e evolução biológica Duração do áudio 5 minutos O objetivo deste programa é tratar das idéias neodarwinistas e mostrar
Teoria sintética da evolução Parte II Conteúdos: Tempo: Objetivos: Idéias evolucionistas e evolução biológica Duração do áudio 5 minutos O objetivo deste programa é tratar das idéias neodarwinistas e mostrar
A situação do câncer no Brasil 1
 A situação do câncer no Brasil 1 Fisiopatologia do câncer 23 Introdução O câncer é responsável por cerca de 13% de todas as causas de óbito no mundo: mais de 7 milhões de pessoas morrem anualmente da
A situação do câncer no Brasil 1 Fisiopatologia do câncer 23 Introdução O câncer é responsável por cerca de 13% de todas as causas de óbito no mundo: mais de 7 milhões de pessoas morrem anualmente da