ASSOCIAÇÕES ENTRE FUNGOS MICORRÍZICOS ARBUSCULARES E TRÊS ESPÉCIES FLORESTAIS DE LEGUMINOSAE EM UMA FLORESTA DE TERRA FIRME NA AMAZÔNIA CENTRAL
|
|
|
- Davi Galindo Santana
- 6 Há anos
- Visualizações:
Transcrição
1 INSTITUTO NACIONAL DE PESQUISAS NA AMAZÔNIA INPA UNIVERSIDADE FEDERAL DO AMAZONAS UFAM Programa de Pós-Graduação em Biologia Tropical e Recursos Naturais PPG/BTRN ASSOCIAÇÕES ENTRE FUNGOS MICORRÍZICOS ARBUSCULARES E TRÊS ESPÉCIES FLORESTAIS DE LEGUMINOSAE EM UMA FLORESTA DE TERRA FIRME NA AMAZÔNIA CENTRAL ALEX BRUNO DOS SANTOS MACIEL Manaus, AM Abril, 2007
2 Livros Grátis Milhares de livros grátis para download.
3 ALEX BRUNO DOS SANTOS MACIEL ASSOCIAÇÕES ENTRE FUNGOS MICORRÍZICOS ARBUSCULARES E TRÊS ESPÉCIES FLORESTAIS DE LEGUMINOSAE EM UMA FLORESTA DE TERRA FIRME NA AMAZÔNIA CENTRAL Orientadora: REGINA C. C. LUIZÃO, Ph.D. Co-orientadora: Dra. CAROLINA VOLKMER DE CASTILHO Fonte Financiadora: Projetos Ecológicos de Longa Duração PELD/Site 1 Dissertação apresentada ao Programa de Pós-Graduação em Biologia Tropical e Recursos Naturais do convênio INPA/UFAM, como parte dos requisitos para obtenção do título de Mestre em CIÊNCIAS DE FLORESTAS TROPICAIS.. Manaus, AM Abril, 2007 ii
4 Maciel, Alex Bruno dos Santos Associações entre fungos micorrízicos arbusculares e três espécies florestais de Leguminosae em uma floresta de terra firme na Amazônia Central/ Alex Bruno dos Santos Maciel Manaus: UFAM/INPA, p. ilust. Dissertação de Mestrado Área de concentração Ciências de Florestas Tropicais. 1. Ecologia Florestal 2. Fungos micorrízicos arbusculares 3. Leguminosae 4. Densidade de esporos 5 Amazônia Central Sinopse: Foram avaliadas as taxas de colonização micorrízica e a densidade de esporos de três espécies arbóreas da família Leguminosae (E. glabriflora, Z. racemosa e A. micrantha) em uma mesoescala (25 Km2) de uma floresta de terra firme na Amazônia Central. Avaliou-se as relações entre as porcentagens de colonização, densidades de esporos com as propriedades físicas e químicas do solo e a contribuição para a nutrição foliar das três espécies de leguminosas quando associadas aos fungos micorrízicos arbusculares. Palavras-chave: ecologia de florestas tropicais, micorrizas arbusculares (MA), associações simbióticas, densidade de esporos, Leguminosae, Amazônia Central. Keywords: Tropical rain forests, arbuscular mycorrhiza (AM), symbiotic associations, spores density, Leguminosae, Central Amazon. iii
5 A Deus, Por me permitir sonhar mais essa vida e evoluir a caminho da luz. À minha mãe Mª Izabela Maciel, Por ser meu anjo na terra me guiando sempre. Ao meu avô Benedito Inácio Maciel (in memorian) Por me ensinar a amar a natureza e ao próximo. À toda minha família Maciel, Por acreditar em mim e me amar incondicionalmente. DEDICO. iv
6 AGRADECIMENTOS À minha orientadora Dra. Regina Celi Costa Luizão por acreditar em mim desde o princípio e pelos conselhos e correções. À minha co-orientadora Dra. Carolina Volkmer de Castilho por ter sido essencial na realização deste estudo com seus conselhos, discussões e por sua dedicação à Reserva Ducke. Ao Dr. Flávio J. Luizão pelo encaminhamento no início dessa odisséia, pela paciência e colaboração em todos os níveis deste estudo. Ao Dr. Pedro M. Antunes (University of Guelph, Ontario, Canadá) por tanto me ajudar por inúmeros s, com incentivo e conselhos na idealização deste estudo. Ao Curso de Pós-Graduação em Ciência de Florestas Tropicais CFT/INPA, nas pessoas do coordenador do curso, Dr. José Francisco e da Ana Lopez e Jaqueline Colaço, que tanto ajudaram. À Capes pela concessão da bolsa de estudo; Ao PELD Site 1, pelo financiamento do meu estudo e apoio logístico para a realização do mesmo. À Coordenação de Pesquisas em Silvicultura Tropical CPST/INPA, em especial ao coordenador e amigo Dr. Paulo de Tarso Sampaio e a Valdecira Azevedo pelo apoio em todos os momentos. Ao Sr. Sebastião Sales(Sabá) por ter sido meus braços e pernas na coleta de todo material no campo e mais ainda por me proteger das jararacas nas trilhas da Ducke. À Rejane Freitas, Cilene Palhêta e Francisco Moreira, imprescindível colaboração na minha iniciação à micorrizologia. À Maria da Conceição Loureiro Campelo (Concita) do Laboratório de Análises de Solos e Plantas da Embrapa/CPAA, por ter sido minha salvadora no momento que tanto precisei. Ao Carlos Moura e ao Rubens (PELD), que me deram total apoio logístico na realização desse estudo, desde o transporte a Reserva Ducke até a aquisição de materiais utilizados neste estudo. Aos companheiros do LTSP/INPA, em especial à Tânia, Edivaldo, Orlando, Jonas, Raimundo, Beto, Morgana, Kleber pelas orientações e apoio nas análises laboratoriais. Aos demais filhos dos Luizão, em especial às minhas irmãs Carol López e Joana de Paula, também a Lucerina, Jean, Ricardo Saci, pela v
7 Fabrício, Veber, Fabiane, Romilda, Erick, pela amizade, apoio nas coletas e momentos agradáveis no laboratório. Aos meus amigos da NATA : Sammy Aquino, Germano Lobo, Penélope Antony, Heleny Ponciano, Rejane Leite e Emanuele Gurgel, pelo companheirismo desde os tempos da faculdade. E a todos os demais companheiros de luta da Engenharia Florestal, principalmente a Evely Bentes, Karol Barbosa, Karina Oliveira, Melissa Chalco. À Amanda Lima pela ajuda com o abstract desse estudo e constante apoio e a Lucinéia Souza pelo incentivo nos trabalhos de pesquisa. Aos meus companheiros GALEROSOS do CFT/INPA: Juliana Peixoto, Matheus Paixão, Rodrigo Serafini, Alexandre Pirani, Mônia Fernandes, Adriana Bariani, Raphael Leduc, Giuliano Guimarães, Gabriel Riva e Cícero Furtado. Também aos da turma de 2004, André, Patrícia, Pauletto, Raquel, Dalva, Renata e os da turma de 2006, Fabiana, Lissandra, Sheron, Heloise e Patrícia, Everton, Pedro. Aos meus amigos de fora do mundo da pesquisa científica, aos que vivem fora da Amazônia e aos de fora do país. Também àqueles que vivem em outro plano e que me protegem todos os momentos. À Reserva Florestal Adolpho Ducke e a todos aqueles que trabalham pela sua conservação e proteção, para que nós pesquisadores possamos continuar estudando-a. vi
8 Artigo 4. Fica decretado que o homem não precisará nunca mais duvidar do homem. Que o homem confiará no homem como a palmeira confia no vento, como o vento confia no ar, como o ar confia no campo azul do céu. Parágrafo único. O homem confiará no homem como um menino confia em outro menino. Artigo final. Fica proibido o uso da palavra liberdade, a qual será suprimida dos dicionários e do pântano do engano das bocas. A partir deste instante a liberdade será algo vivo e transparente como um fogo ou um rio, e sua morada será sempre o coração do homem. Thiago de Mello Estatutos do Homem vii
9 RESUMO Aprofundar o conhecimento sobre as associações micorrízicas arbusculares (MA) pode ser uma importante contribuição para o planejamento de ações visando a conservação e produção de espécies florestais na Amazônia. Este estudo teve como objetivo avaliar a associação entre micorrizas arbusculares (MA) em três espécies arbóreas da família Leguminosae (Eperua glabriflora, Zygia racemosa e Andira micrantha) distribuídas em uma mesoescala (25 km2) de floresta de terra firme na Amazônia Central. O estudo foi conduzido nas 30 parcelas terrestres permanentes (0,25 x 250 m2) da Reserva Florestal Adolpho Ducke, a 26 km ao norte de Manaus, Amazonas. Em junho e julho de 2006 foram coletadas amostras de raízes, solos da rizosfera e folhas maduras das três espécies-alvo, para estimar a colonização micorrízica no comprimento de suas raízes, densidade de esporos nas suas rizosferas e as concentrações de nutrientes presentes nas suas folhas e, além disso, avaliar como estas variáveis são influenciadas pelas propriedades físicas e químicas do solo onde ocorrem os indivíduos. Análises de variância (ANOVA) e teste de Tukey (5%) foram utilizadas para comparação das médias de colonização e número de esporos. Para avaliar as relações entre as variáveis testadas, correlações e regressões lineares também foram realizadas entre as porcentagens de colonização, densidade de esporos, textura do solo e conteúdo de nutrientes nas folhas. Duas espécies, E. glabriflora e A. micrantha, foram fortemente associadas às taxas de colonização micorrízicas. Em Z. racemosa as baixas taxas de colonização micorrízica em suas raízes foram relacionadas negativamente com o Fe2+ presente no solo e em A. micrantha elas foram relacionadas positivamente com Ca2+. As densidades de esporos nas rizosferas responderam diferentemente às propriedades físicas e químicas do solo. Em E. glabriflora a densidade de esporos variou com ph do solo das parcelas. Em Z. racemosa, a quantidade de esporos variou com as concentrações de três macronutrientes no solo (P, Ca2+ e Mg2+) e, em A. micrantha foi a variação da altitude, através da textura do solo, que influenciou as densidades de esporos FMA. A ausência de maiores relações entre a colonização micorrízica e as variáveis edáficas indicam que embora as plantas hospedeiras sejam sensíveis à mudanças na fertilidade do solo e na topografia, os organismos parceiros envolvidos nesta simbiose não necessariamente seguem os mesmos padrões. A diversidade dos fungos micorrízicos que colonizam estas espécies de Leguminosae parece ser específica, uma vez que respondem diferentemente às variáveis ambientais testadas. A identificação taxonômica dos fungos micorrízicos que estão colonizando E. glabriflora, Z. racemosa e A. micrantha poderá confirmar esta tendência. viii
10 ABSTRACT Increasing knowledge on the arbuscular mycorrhizal association (AM) may be an important contribution for planning actions envisaging conservation and production of forest species in the Amazon. The aim of this study is to evaluate the association among arbuscular mycorriza (AM) in three woody species belonging to the Leguminosae family (Eperua glabriflora, Zygia racemosa and Andira micrantha) distributed in a mesoscale (25 km2) of terra-firme forest in Central Amazon. The study was carried out in the 30 permanent plots (0.25 x 250 m2) of the Reserva Florestal Adolpho Ducke, 26 km north of Manaus, Amazonas State. During June and July, 2006 root samples, rizosphere soils, and leaves from the three target species were collected to evaluate the mychorrizal colonization in their roots, rizospheres spores density, nutrient content in their leaves and to evaluate how they are influenced by the physical and chemical soil properties in each individual. Analysis of variance (ANOVA) and Tukey test (5%) were used to compare the average colonization and the number of spores. Linear regression and correlations were also used to evaluate the relationship among colonization percentages, spores density, soil texture and nutrient contents in the leaves. Two species, E. glabriflora and A. micrantha), were strongly associated with mycorrhizal. In Z. racemosa the low micorrhizal colonization rates of the roots were negatively related to Fe2+ in the soil and in A. micrantha they were positively related to Ca2+. The spores density in the rizosphere showed different responses to the soil physical and chemical properties. Spores density E. glabriflora varied with the soil ph. In Z. racemosa, the numbers of spores were related to the concentrations of three macronutrients in the soil (P, Ca2+ e Mg2+) and, in A. micrantha the topography through the soil texture showed to influence the spores density. The absence of major relationship among soil properties and the mychorrizal colonization seems to indicate that though the host plants are sensitive to changes in soil fertility and topography, the partners organisms of this symbiosis not necessarily follow the same patterns. The diversity of mycorrizal fungi colonizing these Leguminosae species seems to be specific, since they responded differently to the environmental variables tested. The taxonomic identification of the mycorrizal fungi that colonize glabriflora, Z. racemosa e A. micrantha may confirm this trend. ix
11 LISTA DE TABELAS Maciel, Alex Bruno dos Santos...iii Sinopse: Foram avaliadas as taxas de colonização micorrízica e a densidade de esporos de três espécies arbóreas da família Leguminosae (E. glabriflora, Z. racemosa e A. micrantha) em uma mesoescala (25 Km2) de uma floresta de terra firme na Amazônia Central. Avaliou-se as relações entre as porcentagens de colonização, densidades de esporos com as propriedades físicas e químicas do solo e a contribuição para a nutrição foliar das três espécies de leguminosas quando associadas aos fungos micorrízicos arbusculares...iii Palavras-chave: ecologia de florestas tropicais, micorrizas arbusculares (MA), associações simbióticas, densidade de esporos, Leguminosae, Amazônia Central...iii Keywords: Tropical rain forests, arbuscular mycorrhiza (AM), symbiotic associations, spores density, Leguminosae, Central Amazon... iii Tabela 1. Correlação de Pearson entre 13 variáveis do solo (de 0 10cm) e 8 nutrientes foliares e três eixos de ordenação produzidos da Análise de Componentes Principais (PCA)...32 Tabela 2. Regressão linear entre as porcentagens de colonização micorrízica PCM nas raízes de E. glabriflora, Z. racemosa e A. micrantha e as características do solo (eixos de PCA)...35 Tabela 3. Concentrações de macro- e micronutrientes presentes em folhas de E. glabriflora, Z. racemosa e A. micrantha. Os números são médias e desvios padrão (entre parênteses)...36 Tabela 4. Regressão linear (r2) da densidade de esporos (DE) em E. glabriflora, Z. racemosa e A. micrantha e os três eixos ordenados de PCA das variáveis do solo...37 Tabela 5. PCM e densidade de esporos (DE) de FMA associados a várias espécies vegetais na Amazônia brasileira Tabela 6. PCM e densidade de esporos (DE) de FMA associados a várias espécies vegetais no Brasil...48 Tabela 7. PCM e densidade de esporos (DE) de FMA associados a várias espécies vegetais em diferentes localidades...49 x
12 LISTA DE FIGURAS Figura 1. Classificação atual das MAs e suas principais características (Modificado do INVAM. Fonte: 13 Figura 2. Imagem de satélite Landsat da Reserva Florestal Adolpho Ducke (RFAD). Fonte: Figura 3. Esquema da disposição das 30 parcelas do PPBio na RFAD. Fonte: (Imagem produzida por D. Drucker) Figura 4. Ilustração das curvas de nível (linha em vermelho) de cada parcela. Fonte: (Imagem produzida por D. Drucker)...21 Figura 5. Ocorrência das espécies de Leguminosae nas 30 parcelas permanentes do PPBio/PELD na RFAD. Trilhas no sentido leste/oeste (LO) numeradas de 3 a 8 e trilhas no sentido norte/sul (NS) numeradas de 1 a 5. Retângulo marcando os locais das parcelas permanentes Figura 6. Distribuição de algumas variáveis físicas e químicas do solo ao longo de um gradiente de altitude de 30 parcelas na RFAD Figura 7. Porcentagem média de colonização micorrízica em seguimentos de raízes de indivíduos de Eperua. glabriflora, Zygia. racemosa e Andira micrantha...34 xi
13 Figura 8. Relação entre a colonização micorrízica (%) (A) nas raízes de Z. racemosa e as concentrações de Fe2+ (mg.kg-1) do solo; (B) nas raízes de A. micrantha e as concentrações de K+ (mg.kg-1)...35 Figura 9. Densidade média de esporos em 10 g de solo coletado na rizosfera de Eperua glabriflora, Zygia racemosa e Andira micrantha Figura 10. Relações entre densidade de esporos presentes na rizosfera de Z. racemosa e os macronutrientes (mg. Kg-1) no solo, Fósforo (A), Cálcio (B) e Magnésio (C)...38 Figura 11. Relação entre densidade de esporos presentes na rizosfera de E. glabriflora e o ph do solo...39 Figura 12. Relações entre densidade de esporos presentes na rizosfera de A. micrantha e a altitude (gráfico A) e % de argila (gráfico B)...39 LISTA DE ABREVIAÇÕES E SIGLAS INPA Instituto Nacional de Pesquisa da Amazônia UFAM Universidade Federal do Amazonas PPG/BTRN - Programa de Pós-Graduação em Biologia Tropical e Recursos Naturais CFT Ciências de Florestas Tropicais PELD Projetos Ecológicos de Longa Duração PPBio Amazônia Programa de Pesquisas em Biodiversidade da Amazônia RFAD Reserva Florestal Adolpho Ducke FMA Fungo micorrízico arbuscular MA Micorriza arbuscular PCM Porcentagem de colonização micorrízica KOH Hidróxido de potássio H2O2 Peróxido de hidrogênio HCl Ácido clorídrico KCl Cloreto de potássio DAP Diâmetro a altura do peito ANOVA Análise de variância xii
14 PCA Análise dos Componentes Principais - SUMÁRIO RESUMO...viii ABSTRACT...ix INTRODUÇÃO...1 OBJETIVOS...5 Geral... 5 Específicos... 5 HIPÓTESES...6 REVISÃO BIBLIOGRÁFICA... 7 A Floresta Amazônica Aspectos Gerais... 7 Solos da Amazônia... 7 Micorrizas Definições Gerais... 8 Classificações das Micorrizas...9 Transferência de Nutrientes entre Fungos Micorrízicos e a Planta Micorrizas Arbusculares Classificação taxonômica dos FMAs Estruturas das MAs Importância das MAs...14 Ecologia das MAs A família Leguminosae xiii
15 Importância econômica da família Associações entre Leguminosae e microrganismos MATERIAL E MÉTODOS Área de estudo Delineamento experimental Seleção das espécies Características das espécies-alvo Amostragem das raízes Colonização micorrízica Amostragem do solo Densidade de esporos Análises químicas e físicas do solo Amostragem e análise química dos tecidos foliares Análise dos dados RESULTADOS...29 DISCUSSÃO...40 CONCLUSÕES...50 REFERÊNCIAS BIBLIOGRÁFICAS APÊNDICE...63 xiv
16 INTRODUÇÃO Os estudos realizados em regiões tropicais são de vital interesse especialmente por duas razões, primeiro porque estes ambientes abrangem imensas áreas e segundo por serem essenciais para os processos biológicos, visto que abriga uma rica biota que interage entre si sob diversas formas (Kellman & Tackaberry, 1997). O bioma Amazônico, reconhecido por ter grande diversidade biótica, possui solos predominantemente com baixa fertilidade química natural e alta acidez (Ranzani, 1980; Sanchez, et al. 1982) estando entre os mais antigos do planeta (Sombroek, 1966), e por milhões de anos em processo de lixiviação, resultando em um déficit nutricional que não supre a necessidade da planta (Oliveira et al., 1997). Para compensar essa restrição, as plantas utilizam mecanismos para suprir suas necessidades nutricionais nesses ambientes. Um dos mecanismos é a interações com microrganismos, para facilitar a captação de nutrientes necessários para seu crescimento, sobrevivência e reprodução. As interações mutualísticas entre variados organismos são importantes em vista do benefício para ambos os organismos envolvidos (Johnson et al., 1997). As interações entre plantas e microrganismos são particularmente importantes porque as plantas representam a principal fonte de carbono, o elemento que limita severamente o crescimento microbiológico no solo. Por outro lado, os microrganismos têm um papel essencial na mineralização de nutrientes em formas disponíveis para absorção pelas plantas (Johnson et al., 2005), na proteção contra patógenos, na diminuição dos impactos de estresses de causas bióticas e abióticas (Smith & Read, 1997), no favorecimento da nodulação e fixação de N2 em leguminosas (Siqueira et al., 2002), no crescimento da planta e agregação do solo (Abbott & Gazey, 1994), dentre outros. A mais comum e possivelmente a mais importante interação mutualística no reino das plantas são as micorrizas, que literalmente significa raiz com fungo. Estas associações simbióticas ocorrem na maioria das plantas vasculares, tanto nativas como cultivadas (Raven et al., 2001). As duas formas de associações micorrízicas mais importantes e comuns são as micorrizas arbusculares (MA), que pertencem ao grupo das endomicorrizas, e as ectomicorrizas (Jones et al., 1998). Entretanto, tem havido
17 uma ênfase maior nos estudos com micorrizas arbusculares, em especial aqueles relacionados a ecossistemas manejados e mais raramente aqueles desenvolvidos em ecossistemas naturais (Moreira & Siqueira, 2002). Com poucas exceções, as micorrizas têm sido observadas em todas as espécies de plantas de interesse econômico (Jackson & Manson, 1984). Essa premissa é confirmada pelos diversos estudos que visam à consolidação das informações sobre as espécies que desenvolvem associações com fungos (Carneiro et al. 1998; Carrenho et al. 2001; Miranda & Miranda, 2001; Johnson et al. 2005). Portanto, é importante aprofundar o conhecimento sobre os padrões de distribuição da associação entre esses fungos e as plantas vasculares em diferentes ambientes florestais. Este entendimento é crucial para o planejamento da conservação das florestas tropicais (Margules et al., 2002; Tuomisto et al., 2003). Entre os elementos que tem a absorção favorecida pelas micorrizas arbusculares, o fósforo se destaca não somente por ser altamente favorecido pela simbiose mas também por ter um papel crucial no desenvolvimento das plantas, pois participa de um grande número de compostos em diversos processos metabólicos. Nas regiões tropicais e subtropicais é um dos elementos que mais limita o crescimento vegetal, pois se encontra em baixa disponibilidade no solo (Eira & Carvalho, 1970; Malavolta, 1980). Em quantidades adequadas, o fósforo estimula o desenvolvimento radicular, é essencial para boa formação de frutos e sementes, e incrementa a precocidade da produção (Raij, 1991). A topografia pode influenciar os estoques e dinâmica dos nutrientes do solo em regiões temperadas e em regiões tropicais, porém esta fonte de variação às vezes tem sido ignorada (Luizão et al., 2004). Muitos autores, citados por Costa et al. (2005), sugerem que o relevo topográfico e os tipos de solo são os maiores determinantes da distribuição de espécies de plantas e padrões de comunidades em pequena escala (1-50 ha) ou em escala regional. A topografia é uma variável que provavelmente reflete outros fatores ambientais e isolada não afeta diretamente a ocorrência de fungos (Claridge et al. 2000). A topografia e a estrutura da vegetação são as variáveis que determinam a distribuição espacial de fungos marasmióides em um período de baixa pluviosidade, possivelmente devido a influência da umidade na liteira (Braga-Neto, 2006). Portanto, no caso da associação fungo micorrízico-planta a
18 topografia relacionada a outras variáveis ambientais também pode ser determinante para o sucesso dessa simbiose. A diversidade e densidade dos propágulos de FMA no solo estão relacionadas indiretamente com as condições ecológicas de cada ecossistema (Maia & Trufem, 1990). Sendo assim, os estudos ecológicos são necessários para o entendimento dos processos que levam ao estabelecimento e manejo populacional dos fungos, com ênfase nas taxas de formação micorrízica e quantidade de propágulos, incluindo os esporos (Abbott & Gazey, 1994). Na região Amazônica os estudos relacionados a associação das micorrizas arbusculares e espécies florestais nativas ainda é muito recente, sendo que a maioria desses trabalhos estão mais relacionados a presença ou ausência da associação nas raízes das plantas (St. John, 1980; Guitton, 1996, Moreira et al., 1997) e poucos são aqueles que relacionam essa associação aos diversos fatores ambientais (Ropero, 2007, Oliveira et al., 1999) e mais escassos ainda aqueles que descrevem a diversidade populacional desses microrganismos simbióticos (Freitas, 2005; Leal, 2005). Oliveira et al. (1999), relatam que apenas a presença do fungo na raiz não significa o favorecimento da planta na absorção de nutrientes dos solos, uma vez que a eficiência da simbiose pode variar, dependendo da planta, do fungo e do ambiente. Os estudos da associação entre os fungos micorrízicos e plantas nativas da Amazônia deveriam ser mais estimulados em vista da necessidade de se entender essas interações e posteriormente multiplicar tais conhecimentos visando à larga produção de essências florestais adaptadas a ambientes degradados. Muitas espécies da família Leguminosae são promissoras para suprir tais necessidades, pois, além de possuírem grande potencial econômico, também desenvolvem um importante papel ecológico em diversos ecossistemas, pois têm a capacidade de formar simbiose com dois microrganismos (rizóbios e fungos micorrízicos), uma importante estratégia para seu estabelecimento em solos com baixos teores de nutrientes (Bonetti et al., 1984; Vasconcelos et al., 1984; Silva et al., 1994). Este estudo visa fornecer informações pertinentes às relações ecológicas da mais comum forma de associação simbiótica, as micorrizas arbusculares (MA), encontradas em três espécies florestais da família Leguminosae (Eperua glabriflora, Zygia racemosa e Andira micrantha) e
19 variáveis ambientais ligadas ao fator altitude topográfica que compõem as florestas tropicais. Tal entendimento é importante para o planejamento de futuros projetos que visem à conservação das florestas e à produção de espécies florestais.
20 OBJETIVOS Geral Avaliar a associação entre fungos micorrízicos arbusculares (FMA) associados a três espécies arbóreas da família Leguminosae (Eperua glabriflora, Zygia racemosa e Andira micrantha) encontradas em uma mesoescala (25 Km2) de floresta de terra firme na Amazônia Central. Específicos (i) Avaliar as diferenças entre a colonização por FMA (%) estabelecidas em raízes de Eperua glabriflora, Zygia racemosa e Andira micrantha distribuídas em um gradiente topográfico de floresta; (ii) Avaliar as relações entre a colonização micorrízica (%) nas raízes e as concentrações de macro- e micronutrientes dos tecidos foliares de Eperua glabriflora, Zygia racemosa e Andira micrantha; (iii) Avaliar as relações entre a colonização micorrízica (%) nas raízes de Eperua glabriflora, Zygia racemosa e Andira micrantha e as propriedades químicas e físicas do solo (nutrientes, ph, umidade e textura); (iv) Avaliar as diferenças entre as densidades de esporos de FMA presentes na rizosfera e as suas relações com as porcentagens de colonização micorrízica em Eperua glabriflora, Zygia racemosa e Andira micrantha distribuídas em um gradiente topográfico de floresta; (v) Avaliar as relações entre a densidade de esporos de FMA nas rizosferas de Eperua gabriflora, Zygia racemosa e Andira micrantha, e as propriedades químicas e físicas do solo (nutrientes, ph, umidade e textura).
21 HIPÓTESES Hipótese 1 Há diferenças significativas entre a colonização por FMA (%) em raízes de Eperua glabriflora, Zygia racemosa e Andira micrantha estabelecidas em um gradiente topográfico de floresta. Hipótese 2 Há relações entre a colonização micorrízica (%) nas raízes e os teores de macro- e micronutrientes dos tecidos foliares de Eperua glabriflora, Zygia racemosa e Andira micrantha. Hipótese 3 Há relações entre a colonização micorrízica (%) nas raízes de Eperua glabriflora, Zygia racemosa e Andira micrantha e as propriedades químicas e físicas do solo (nutrientes, ph, umidade e textura). Hipótese 4 Há diferenças entre as densidades de esporos de FMA presentes na rizosfera e as suas relações com as porcentagens de colonização micorrízica em Eperua glabriflora, Zygia racemosa e Andira micrantha distribuídas em um gradiente topográfico de floresta. Hipótese 5 Há relações entre a densidade de esporos de FMA nas rizosferas de Eperua gabriflora, Zygia racemosa e Andira micrantha, e as propriedades químicas e físicas do solo (nutrientes, ph, umidade e textura).
22 REVISÃO BIBLIOGRÁFICA A Floresta Amazônica Aspectos Gerais A floresta amazônica, com aproximadamente 7 milhões de km2, é caracterizada por uma alta diversidade biológica, mas pouco se sabe ainda sobre as espécies que a compõem e suas relações filogenéticas. As espécies vegetais nativas da floresta Amazônica são ainda pouco conhecidas. Até hoje, aproximadamente 20 espécies são mundialmente conhecidas e utilizadas por madeireiros, e apenas poucas são listadas como frutíferas ou por suas propriedades medicinais (Ribeiro et al., 1999). O modelo de ocupação tradicional ao qual a Amazônia tem sido submetida tem levado ao aumento significativo do desmatamento na Amazônia Legal, sendo este um fenômeno de natureza bastante complexa, que não pode ser atribuído a um único fator (Alencar et al., 2004). As atividades econômicas que exploram em larga escala os recursos da Amazônia Legal brasileira têm aumentado drasticamente a taxa de desmatamento que, no período de 2004 e 2005, foi registrada à marca de km2 (Inpe, 2007). Recentemente, a criação de novas políticas públicas, fundamentadas no desenvolvimento sustentável da região, têm ajudado na conscientização local para a diminuição do desmatamento desnecessário de áreas. A Amazônia é hoje uma região em si, que demanda uma política de consolidação do desenvolvimento. Fortalecimento institucional, C/T&I e regionalização são estratégias sugeridas para esse fim (Becker, 2005). Solos da Amazônia Há dois tipos de solos predominantes na região Amazônica: os latossolos (oxissolos) e os podzóls (ultissolos), que representam cerca de 75% do total (Sanchez et al., 1982). Os latossolos são profundos, de boa drenagem e estrutura, baixo ph, alta saturação de alumínio e baixos teores de fósforo, potássio, cálcio, magnésio, zinco e outros micronutrientes. Os podzólicos são solos distróficos que possuem horizonte B argílico, apresentando características físicas menos favoráveis, destacando-se aqui os podzólicos vermelho-amarelo distróficos e a terra roxa estruturada distrófica (Kitamura, 1994).
23 Os nutrientes necessários para as plantas encontram-se em uma pequena fração (menor que 0,2%) que está dissolvida na solução do solo. Aproximadamente 98% dos bioelementos no solo estão na forma de liteira, húmus, ligados ao material inorgânico de difícil solubilização ou ainda incorporados aos minerais. Essa grande porção atua como uma reserva, a qual, por meio da decomposição e mineralização, disponibiliza lentamente os nutrientes para o solo. Os 2% restantes estão dissolvidos nos colóides do solo. Entre a solução do solo, os colóides do solo e a reserva mineral há um equilíbrio dinâmico que disponibiliza continuamente um suprimento de nutrientes para as plantas (Larcher, 2000). A floresta densa ocorre mesmo em solos quimicamente pobres, pois sua estratégia nutricional vem dos nutrientes contidos na cobertura vegetal, cujas partes mortas, ao caírem ao solo, são mineralizadas e recicladas (Pires, 1978). Além disso, a vegetação presente nestes solos passou por um processo de adaptação, utilizando eficientemente os nutrientes reciclados da matéria orgânica decomposta pelos microrganismos dos solos (Oliveira et al., 1997). Micorrizas Definições Gerais Na luta pela sobrevivência, fungos e plantas desenvolveram a capacidade de se comunicarem molecularmente, através de mecanismos de reconhecimento, tropismo e tactismo, que permitiram a interação célula-célula e a integração morfológica e funcional, que são requisitos essenciais à formação de uma simbiose mutualística, como ocorre entre plantas e fungos micorrízicos (Moreira & Siqueira, 2002). Micorriza (mykes = fungo; rhiza = raiz) é uma associação simbiótica que ocorre entre fungos do solo e raízes da maioria das plantas vasculares (Sieverding, 1991). A associação é caracterizada pela formação de um grupo funcional, através do qual ocorre o movimento de fotoassimilados da planta para o fungo e de nutrientes absorvidos do solo pelo fungo para a planta. Embora tenha surgido há mais de 400 milhões de anos, quando as plantas iniciaram o processo de colonização do habitat terrestre, as micorrizas só foram reconhecidas e tratadas cientificamente em meados do século XIX, quando foram publicados os primeiros relatos detalhados da associação entre células radiculares e micélios fúngicos (Siqueira, 1986).
24 Os fungos micorrízicos colonizam as células de uma raiz viva, principalmente células corticais ou epidérmicas e contribuem aumentando o seu volume no solo e conseqüentemente aumentando a habilidade da planta na absorção da água e dos elementos essenciais, especialmente fósforo (Raven et al. 2001; Tinker, 1975; Islam & Ayanaba, 1981). O aumento na taxa de absorção de P para plantas pode ser de 3 a 5 vezes (Schachtman et al., 1998). Incrementos na absorção de Zn, Mn e Cu, três outros elementos essenciais, têm sido igualmente demonstrados (Raven et al. 2001). Oliveira et al. (1999) verificaram correlações positivas entre a colonização radicular e os teores de Ca, Mg, P e K no tecido foliar de oito espécies florestais da Amazônia. A simbiose fungo/planta não só aumenta a biomassa vegetal, conseqüência da maior absorção de nutrientes, como também influencia a proporção na qual esta se distribui entre a parte aérea e a raiz. O estímulo da absorção de nutrientes e posterior translocação destes à parte aérea causa, relativamente, menor transferência de fotossintatos à raiz e maior retenção na parte aérea, sendo utilizados na produção de matéria verde. Conseqüentemente, a relação peso da matéria seca da parte aérea/peso da matéria seca da raiz é, em geral, mais alta em plantas micorrizadas (Silveira, 1992). Classificações das Micorrizas As micorrizas são classificadas de acordo com o arranjo das hifas nas células do córtex da raiz (Krügner & Tomazello Filho, 1981). Essa classificação das micorrizas em grupos e tipos pode variar bastante de autor para autor (Moreira & Siqueira, 2002). Segundo Brundrett et al. (1996), existem pelo menos sete tipos diferentes de associações micorrízicas (que se agrupam em ectomicorrizas e endomicorrizas), envolvendo diferentes grupos taxonômicos de fungos e plantas e diferentes padrões de alterações morfológicas que ocorrem durante o estabelecimento da colonização pelo fungo. A maioria dos fungos micorrízicos forma ectomicorrizas. São mais de 4000 espécies, que pertencem predominantemente às classes Ascomicetos e Basidiomicetos (Brundrett et al., 1996). Nas ectomicorrizas, a raiz do
25 hospedeiro é recoberta externamente por um manto espesso de hifas, e a penetração do micélio interno no córtex da raiz é sempre intercelular, formando a chamada rede de Hartig (Silveira, 1992). As endomicorrizas caracterizam-se pela penetração da parede das células do córtex radicular pelas hifas do fungo; nas ectomicorrizas, o fungo não penetra a parede celular, porém forma um manto que reveste toda a área do córtex da planta. Nas ectendomicorrizas, quando o fungo penetra as células da planta hospedeira formando ao mesmo tempo um manto. Essas categorias contudo, podem ser representações inadequadas para descrever o complexo conjunto de interações entre micorrizas (Allen, 1991). As endomicorrizas são representadas por três tipos distintos: (a) Ericóide micorriza formada por espécies vegetais dos gêneros Calluna, Rododendro, Erica e Vaccinium da família Ericaceae. As células corticais e epidérmicas das raízes muito finas são invadidas quase que totalmente pela hifa fúngica, formando pelotões (Englander, 1982); (b) Orquidóide considerada como uma das mais complexas interações simbióticas, onde o balanço dinâmico entre os simbiontes permite que o fungo invada progressivamente o tecido da planta, enquanto esta faz uso dos metabólitos fúngicos, de forma parcial ou total. Uma quebra nesse balanço acarreta a eliminação da micorriza ou a mudança da relação para parasitismo. As plântulas de orquídeas são totalmente dependentes do fungo micorrízico, que, invadindo o córtex da raiz, intracelularmente, fornece carboidratos e minerais à planta. Os fungos envolvidos são principalmente do gênero Rhizoctonia e basiomicetos, tais como Armillaria, Marasmius e Fomes (Warcup & Talbot, 1980), e (c) Micorriza Arbuscular (MA) de ocorrência praticamente universal nas mais variadas ordens e famílias de plantas superiores e inferiores, de forma generalizada no ambiente terrestre (Smith & Read, 1997). A ectendomicorriza é uma forma de transição entre a ecto e a endomicorriza. As raízes da planta hospedeira são recobertas externamente pelo manto de hifas, que pode ser reduzido ou mesmo ausente; a rede de Hartig é bem desenvolvida e a penetração do micélio é intra e intercelular. A micorriza Monotropóide possui um manto fúngico bem desenvolvido e ocorre em membros aclorofilados da sub-família Monotropoideae da família Ericaceae, envolvendo basidiomicetos do gênero Boletus. A micorriza Arbutóide, por sua vez, possui manto, hifa externa e uma rede de Hartig bem desenvolvida, sendo
26 que a penetração intracelular forma intensa infecção na forma de pelotões. Ocorre em espécies da família Pyrolaceae e Ericaceae, tendo como microssimbiontes, fungos que, em geral, formam ectomicorrizas como Amanita, Cortinarius e Boletus (Harley & Smith, 1983). Transferência de Nutrientes entre Fungos Micorrízicos e a Planta Pouco se sabe sobre os mecanismos de transferência dos nutrientes minerais absorvidos pelos fungos micorrízicos às células das raízes dos vegetais. Com as ectomicorrizas, os fosfatos inorgânicos podem simplesmente difundir-se das hifas na rede de Hartig e ser absorvidos pelas células corticais das raízes. Com as endomicorrizas, o processo pode ser mais complexo. Os nutrientes podem se difundir de arbúsculos intactos para as células corticais. Simultaneamente, arbúsculos senescentes podem liberar seus conteúdos internos para as células radiculares do hospedeiro (Taiz & Zeiger, 2004). Outro fator chave na determinação da extensão da associação micorrízica com a raiz da planta é o estado nutricional da hospedeira. A deficiência moderada de um nutriente com o fósforo tende a promover a infecção, enquanto plantas com nutrientes abundantes tendem a suprimir a infecção micorrízica (Taiz & Zeiger, 2004). Micorrizas Arbusculares As micorrizas arbusculares (MA) são associações formadas entre fungos pertencentes à Ordem Glomerales da Divisão Glomeromycota, os quais formam estruturas como arbúsculos, hifas e vesículas no interior do córtex radicular. Esses fungos colonizam as raízes de plantas de quase todos os gêneros das Gimnospermas e Angiospermas, além de algumas representantes das Briófitas e Pteridófitas (Trappe, 1987). As associações micorrízicas arbusculares ocorrem em 83% das dicotiledôneas, 79% das monocotiledêneas e praticamente em todas as gimnospermas (Wilcox, 1990). As MAs podem ser encontradas em plantas herbáceas, arbustivas ou arbóreas que ocupam os mais diversos ecossistemas, como florestas, desertos, dunas, savanas, campos e agrossistemas. O longo tempo de coevolução entre as FMAs e as plantas pode explicar a presença ubíqua desses
27 fungos nos ecossistemas. Eles invariavelmente associam-se à maioria das plantas nativas dos trópicos e a espécies importantes economicamente e/ou ecologicamente (Siqueira et al., 2002). Classificação taxonômica dos FMAs A classificação dos FMAs vem sendo definida ao longo das últimas décadas pela congruência de aspectos morfológicos e moleculares. Atualmente, estes fungos se encontram classificados na Ordem Glomerales da recém proposta Divisão Glomeromycota (Schussler et al., 2001). Eles são considerados um grupo monofilético, ou seja, um grupo de espécies derivadas de um ancestral comum, contendo todos os organismos que formam associações mutualísticas com as raízes das plantas caracterizadas pela formação de arbúsculos (Morton, 1993). Segundo esta classificação, a ordem Glomerales contém duas sub-ordens, Glominaceae e Gigasporineae. A sub-ordem Gigasporineae é formada por uma família, Gigasporaceae, que contém os gêneros Gigaspora e Scutellospora cujos organismos produzem esporos numa célula bulbo suspensora, formam células auxiliares, e possuem uma camada permanente envolvendo a camada laminar na parede dos esporos. A sub-ordem Glomineae é formada por duas famílias, Glomaceae, que contém o gênero Glomus, e a família Acaulosporaceae, com os gêneros Acaulospora e Entrophospora. Os fungos da subordem Glomineae formam hifas cilíndricas com ramificações perpendiculares, formam vesículas dentro das raízes, e os esporos possuem uma ou mais camadas evanescentes envolvendo a camada laminar da parede. Recentemente, Morton & Redcker (2001) propuseram duas novas famílias com dois gêneros respectivos, Archaeosporaceae (gênero Archaeospora) e Paraglomaceae (gênero Paraglomus). Na família Archaeosporaceae, ocorre a formação de um sáculo esporífero prioritário ao desenvolvimento dos esporos na lateral (em Acaulospora) ou dentro (em Entrophospora) da hifa suspensora. Nas duas novas famílias, os esporos são formados em talo da hifa esporógena (Archaespora) ou como em Glomus na Paraglomaceae. Em Gisgasporaceae, os esporos são formados individualmente a partir de uma célula bulbo, por brotação, conforme ilustrados no esquema da Figura 1.
 Estruturas das MAs As MAs possuem três componentes: a raiz da planta hospedeira, as estruturas formadas no córtex radicular (arbúsculos e vesículas) e o micélio e esporos extra-radiculares.")
28 Figura 1. Classificação atual das MAs e suas principais características (Modificado do INVAM. Fonte: Estruturas das MAs As MAs possuem três componentes: a raiz da planta hospedeira, as estruturas formadas no córtex radicular (arbúsculos e vesículas) e o micélio e esporos extra-radiculares. As vesículas, estruturas globosas ou alongadas contendo grânulos de glicogênio e lipídios, são consideradas estruturas de estocagem dos fungos e podem ser formadas dentro ou fora das células do córtex. Os arbúsculos são as estruturas características das MAs. São formados pela intensa ramificação de hifas intracelulares e são responsáveis pela troca de nutrientes entre os simbiontes. O desenvolvimento de arbúsculos é acompanhado da invaginação da membrana plasmática vegetal, não comprometendo a integridade das células radiculares. As hifas intra e extraradiculares são importantes como propágulos para iniciar nova colonização, para gerar novos esporos, para aquisição de nutrientes e ainda podem favorecer a agregação do solo. O micélio extra-radicular pode atingir de 14 a 50 m g-1 de solo, diferenciando-se em hifas absortivas, as quais formam uma rede dicotomicamente ramificada que se estende solo adentro para absorção de nutrientes e água (Siqueira et al., 2002). Os esporos formados pelos FMAs são assexuados e servem para a sua disseminação e sobrevivência. Possuem diâmetro que varia de 45 a 700 µm, coloração hialina, amarelada, esverdeada, amarronzada ou mesmo preta, e
29 forma globosa, alongada ou muitas vezes irregular (Morton, 1988), podendo ter parede lisa ou ornamentada. Os tipos de esporos são distinguíveis em função de sua ontogenia, que justamente com características estruturais, formam a base da taxonomia e sistemática dos FMAs. Apesar dos esporos germinarem facilmente em meios de cultura, o crescimento continuado e a esporulação dos FMAs não ocorrem na ausência de células vivas do hospedeiro. As dificuldades de se cultivar os FMAs in vitro representam um obstáculo ainda não superado, o qual tem limitado os estudos de sua biologia e desenvolvimento biotecnológico (Siqueira et al., 2002). Importância das MAs A associação micorrízica é uma estratégia desenvolvida pelos FMAs, ao longo de milhões de anos, para enfrentar os estresses bióticos sofridos pelas plantas e, por isso, contribui para a sobrevivência, crescimento e produtividade das comunidades vegetais. Segundo Siqueira (1994), as MAs são de interesse especial para as regiões tropicais e subtropicais, especialmente para o Brasil, devido: a) à baixa fertilidade dos solos e elevado requerimento de nutrientes para a maioria das culturas, b) a condições ambientais estressantes, c) à alta proporção de minifundiários com pequeno poder aquisitivo, d) ao suprimento limitado de fertilizantes em certas áreas, e) alto custo de financiamento agrícola, f) à possível exaustão dos depósitos de fosfatos e g) à preocupação crescente com qualidade ambiental, visando minimizar os impactos da poluição do solo e água e do desmatamento. Assim, as MAs são componentes importantes dos ecossistemas e desempenham papel crucial para sua funcionalidade e sustentabilidade. Ecologia das MAs A formação de micorrizas resulta de uma interação complexa de três elementos: o fungo, a planta e o ambiente. Conseqüentemente, diversos fatores ligados a esta trilogia afetam de certa forma direta ou indiretamente o processo de colonização (Bellei & Carvalho, 1992). Assim, o conhecimento das relações ecológicas nos sistemas micorrízicos é essencial para se avaliar a atividade destes fungos e sua simbiose como um componente funcional dos ecossistemas (Couto, 2000).
30 As MAs são importantes componentes dos ecossistemas, com forte influência sobre a composição da comunidade vegetal e sobre a sua funcionalidade (Bever et al., 2002; Hart et al., 2001). Ao mesmo tempo, a ocorrência e distribuição dos FMAs são influenciadas por diversos fatores ambientais (Moreira & Siqueira, 2002). Dentre os fatores bióticos, destacam-se as espécies de plantas, cobertura vegetal, sistema radicular, nutrição vegetal, ciclo e taxa de crescimento, sistemas de cultivo, alelopatia, exsudação de resina e organismos do solo. Quanto aos fatores abióticos, os principais são a disponibilidade de nutrientes, ph (Lohman, 1927), elementos tóxicos (Al, Mn e outros), salinidade, textura, estrutura e agregação do solo, umidade, intensidade luminosa, temperatura, precipitação, poluição atmosférica e do solo, fertilizantes e corretivos, biocidas e práticas culturais e, ainda, contaminação por metais pesados. A estabilidade de ecossistemas naturais, tendo a presença constante de hospedeiros e ausência de variações bruscas na fertilidade do solo, pode garantir a sobrevivência de algumas espécies de FMAs (Siqueira et al., 1989). A ausência de vegetação e a erosão acentuada do solo reduzem ou até mesmo eliminam as MAs do sistema. Isso se deve ao fato dos FMAs serem biotróficos obrigatórios e por isso dependem do C da fotossíntese (raízes) para seu desenvolvimento (Moreira & Siqueira, 2002). Para Silveira (1992), diferentes tipos de interações podem ocorrer entre fungos micorrízicos e outras populações microbianas da rizosfera. Alguns componentes da microbiota do solo como fungos, bactérias e actinomicetos, podem atuar como antagonistas e comensalistas em relação aos fungos micorrízicos. Também a presença de nematóides micófagos pode reduzir o micélio e o número de esporos. A fauna do solo tem participação ativa sobre os fungos micorrízicos, atuando na dispersão de propágulos e alimentando-se de hifas, influenciando então no estabelecimento dos fungos micorrízicos no solo. A luminosidade é uma outra variável que afeta a colonização, uma vez que plantas cultivadas em alta luminosidade absorvem íons mais rapidamente do que aquelas mantidas em baixa irradiância, pois estas últimas alocam menos fotoassimilados para as raízes. Os açúcares translocados para as raízes são utilizados na produção de energia (ATP) e como substrato para o crescimento do sistema radicular, o que aumenta o volume de solo explorado.
31 A disponibilidade de energia é essencial para a absorção de íons (Marenco & Lopes, 2005). Guitton (1996) observou a influência da sazonalidade nas colonizações por FMA em espécies florestais como cumarú, jacareúba e marupá nativas da Amazônia e observou variação significativa nos percentuais médios de colonização. Moreira & Siqueira (2002) descrevem as características das plantas que afetam a simbiose e destacam a condição micorrízica, o estado nutricional local, o ciclo e taxa de crescimento e produção de substâncias alelopáticas. Segundo os autores, as plantas são bem conhecidas pela capacidade de produzirem uma enorme variedade de substâncias bioativas como os aleloquímicos, cujos efeitos sobre as micorrizas ainda não são bem conhecidos. Certos compostos aleloquímicos podem atuar como inibidores ou estimulantes da micorrização e interferir na ocorrência da simbiose. A família Leguminosae A família Leguminosae representa um dos principais e mais importantes grupos de plantas dentre as fanerógamas e, quantitativamente, está entre as três maiores famílias botânicas, juntamente com a Orchidaceae e a Asteraceae. Ainda assim, a sua extensão, especialmente no que se refere ao número de espécies que constituem a família e a sua distribuição geográfica, diverge entre os autores: Allen & Allen (1981) estimam a família com 748 gêneros, abrigando cerca de espécies, enquanto Corby (1981) considera que as leguminosas abrangem cerca de 649 gêneros e um total de espécies. Suas espécies estão representadas nos trópicos úmidos, nas regiões temperadas, nas zonas áridas, na vegetação de altitude ou montanhosa, nas savanas e nas terras baixas inundadas, como igapós, as várzeas e as florestas de terra firme da região amazônica. Na natureza, as espécies de Leguminosae assumem os mais diversificados tipos de hábitos ou formas vegetativas de vida, desde árvores de pequeno a grande porte, arbustos, cipós e ervas que ocorrem nos mais variados habitats, incluindo ambientes bastante antagônicos como meios aquáticos, as várzeas e igapós da Amazônia e ainda, ambientes xerófitos, como savanas, campinas e campinaranas amazônicas (Allen & Allen, 1981).
32 A América tropical é o maior centro de diversidade de Leguminosae e o Brasil é, particularmente, muito rico em espécies silvestres da família. Na Amazônia, as leguminosas estão representadas por cerca de 148 gêneros, o que corresponde a quase 1/3 do total da família (3.100 taxa específicos e intraespecíficos), com padrão de distribuição disjunta e/ou contínua e com cerca de 21 gêneros provavelmente endêmicos (Allen & Allen, 1981). Importância econômica da família A família Leguminosae é uma das mais importantes famílias botânicas por ser fonte de produtos alimentares, medicinais, ornamentais, madeireiros, além de outros produtos de grande valor econômico (Di Stasi & Hiruma-Lima, 2002). Muitas espécies de Leguminosae são promissoras comercialmente, pois são fontes de produtos como proteínas, óleos, corantes, resinas utilizadas para produzir vernizes, tintas e lacas, além de suas flores fornecerem base para um mel de excelente qualidade. Entre os gêneros mais conhecidos estão a Acacia, Andira, Bauhinia, Caesalpinia, Copaifera, Dalbergia, Dipteryx, Hymenaea, Inga, Mimosa, Phaseolus, Senna, etc. (Lorenzi, 1992). Na Amazônia mais de espécies de leguminosas são árvores de grande porte produtoras de madeira, e freqüentam permanentemente a lista de espécies que são exportadoras por seu valor madeireiro, tais como o Angelim vermelho (Dinizia excelsa), o jacarandá do Pará (Dalbergia paraense) e a cedrorana (Cedrelinga catenaeformis), por exemplo (Souza, 2006). Entre os principais produtos obtidos das leguminosas arbóreas da Amazônia, pode-se citar o óleo-de-copaíba (extraído de perfurações no tronco de árvores de Copaifera multijuga, a copaíba), o verniz copal (extraído de Hymeneae courbaril, o jatobá, cujos frutos também têm alto valor alimentício, contendo uma polpa farinácea bastante apreciada pelas populações da floresta). Há também árvores de cumarú (Dipteryx odorata), que possuem as sementes com alto conteúdo de cumarina, utilizada na indústria de cosméticos (Souza, 2006). Adicionalmente, a elevada biodiversidade natural da vegetação amazônica inclui inúmeras espécies com potencial de exploração por seus princípios curativos e medicinais Dimorphandra mollis, a fava d anta; Crudia amazonica, a orelha de cachorro, e a própria copaíba Copaifera multijuga, e
33 também espécies com potencial para produção de gomas naturais (como Parkia discolor, a faveira-do-igapó, que produz grande quantidade de goma no fruto, com propriedades similares às da goma arábica), e outros (Souza, 2006). Esta família classifica-se ainda como a segunda mais importante na produção de sementes alimentícias, ricas em proteínas e carboidratos, essenciais na dieta humana e animal (Beltrati, 1994). Associações entre Leguminosae e microrganismos Plantas da família Leguminosae podem formar associações simbióticas tanto com bactérias fixadoras de nitrogênio, chamadas de rizóbio, como com fungos micorrízicos arbusculares. Nessa associação mutualista tripartida, os fungos micorrízicos podem aumentar a solubilização e a absorção de fósforo do solo através do micélio extra-radicular, translocando esse elemento para o vegetal. Por sua vez, o rizóbio, localizado em células especiais do nódulo, fixa o nitrogênio atmosférico disponibilizando-o sob forma de amônia para a planta, a qual, em contrapartida, fornece carboidratos para ambos os microssimbiontes (Allen & Allen, 1981; Harley & Smith, 1983, Gross et al., 2004)

34 MATERIAL E MÉTODOS Área de estudo Este estudo foi realizado na Reserva Florestal Adolpho Ducke (RFAD), uma área de 100km2 localizada a 26 km ao norte da cidade de Manaus/AM (Figura 2). Na RFAD, a precipitação varia de a mm por ano. A topografia da reserva é constituída de áreas de platôs e baixios, onde ocorrem pequenos riachos em duas redes de drenagem a leste e oeste da Reserva. Na rede oeste, as nascentes originam uma seqüência de igarapés que desaguam no rio Negro, e na rede leste eles seguem para o rio Amazonas. A altitude varia entre aproximadamente 40 e 110 metros acima do nível do mar. Figura 2. Imagem de satélite Landsat da Reserva Florestal Adolpho Ducke (RFAD). Fonte: A RFAD é coberta por floresta tropical úmida de terra-firme, com altura média de dossel entre 30 e 35 metros. A estrutura e a florística são definidas principalmente pelo tipo de solo e relevo: platô, vertente, campinarana e baixio. Nos platôs os solos são argilosos (latossolo amarelo-álico), sedimentos mais antigos, e nas partes mais baixas arenosas (podzóis e areias quartzosas). A floresta de vertente é de difícil delimitação ocorrendo em solos areno-argilosos.
.")
35 A vertente corresponde a um gradiente fisionomicamente mais similar ao platô nas partes mais altas onde o solo é argiloso e com campinarana nas partes mais baixas onde o solo é areno-argiloso (Ribeiro et al., 1999). Delineamento experimental A RFAD tem um sistema de trilhas distribuídas sistematicamente por uma área de 64 km2, consistindo numa grade de 18 trilhas de 8 km cada, nove trilhas na direção norte-sul e nove trilhas na direção leste-oeste, formando parcelas de 1 km2. Todas as trilhas têm cota de nível altimétrico marcadas a cada 100m, com placas de alumínio fixadas em tubo de PVC de 50 cm de altura. Ao longo do sistema de trilhas estão demarcadas 72 parcelas terrestres permanentes, criadas pelo Projetos Ecológicos de Longa Duração (PELD/CNPq), que possibilitam o estudo integrado e o monitoramento da distribuição de organismos de diferentes grupos biológicos nessa mesoescala. Para este estudo foram utilizadas as 30 parcelas (Figura 3) inseridas no Programa de Pesquisas em Biodiversidade da Amazônia (PPBio Amazônia/ MCT). Figura 3. Esquema da disposição das 30 parcelas do PPBio na RFAD. Fonte: (Imagem produzida por D. Drucker).
36 As parcelas terrestres permanentes são longas e estreitas, com seu maior eixo orientado ao longo das curvas de nível do terreno. Esse desenho minimiza a variação interna de topografia e solo em cada parcela, e permite o uso destas variáveis como preditoras das distribuições de espécies. Cada uma das 30 parcelas tem área de 1 ha (40m x 250m) e segue a curva de nível do terreno (isoclina) (Magnusson et al., 2005) (Figura 4). As coletas do material para este estudo foram realizadas durante os meses de junho e julho de 2006 nas 30 parcelas permanentes do PPBio Amazônia, representando o gradiente topográfico encontrado na RFAD. Esse período representa a transição entre as estações chuvosa e seca na região. Cada parcela representa uma unidade amostral. Figura 4. Ilustração das curvas de nível (linha em vermelho) de cada parcela. Fonte: (Imagem produzida por D. Drucker).
37 A altitude do terreno em todas as parcelas foi medida por um topógrafo profissional. O valor de altitude de cada parcela corresponde à cota de altitude da trilha no ponto de início da parcela. Como as parcelas seguem uma curva de nível, o valor de altitude é constante em toda extensão da parcela. Seleção das espécies A seleção das espécies para este estudo foi realizada previamente através de um levantamento das espécies mais abundantes de Leguminosae encontradas nas parcelas do Projeto PPBio Amazônia (Castilho, dados não publicados). O levantamento mostrou a ampla distribuição da família Leguminosae em todas as parcelas, sendo as espécies Eperua grabriflora, Zygia racemosa e Andira micrantha as mais abundantes. Procedeu-se então a seleção de indivíduos dessas espécies com DAP entre 10-30cm. Portanto, as amostras utilizadas nesse estudo foram coletadas de parcelas que continham indivíduos suficientes para compor uma amostra (com n variável), resultando assim em 141 árvores amostradas (Figura 5). Em cada indivíduo foram coletadas amostras de raízes para análise da colonização micorrízica, de solo para análise das propriedades físicas e químicas e contagem dos esporos e de folhas para análise dos macro- e micronutrientes em cada indivíduo.
38 Figura 5. Ocorrência das espécies de Leguminosae nas 30 parcelas permanentes do PPBio/PELD na RFAD. Trilhas no sentido leste/oeste (LO) numeradas de 3 a 8 e trilhas no sentido norte/sul (NS) numeradas de 1 a 5. Retângulo marcando os locais das parcelas permanentes. Características das espécies-alvo Eperua glabriflora (Ducke) Cowan Sinônimo: Eperua bijuga Benth. var. glabriflora Ducke. Árvore encontrada freqüentemente em áreas de platô, vertente e campinaranas da Amazônia Central e Oriental, pertencente à família Leguminosae: Caesalpinioideae. É popularmente conhecida como muirapiranga e muirapiranga da folha grande. Possui o tronco pouco achatado com fissuras finas. Ritidoma marrom. Folíolos glabros, coriáceos, com aspecto reticulado na face abaxial. Seus folíolos são facilmente reconhecíveis na liteira da floresta por sua margem revoluta (Ribeiro et al., 1999). Sua importância econômica está no uso de sua madeira. Zygia racemosa (Ducke) Barneby & Grimes - Sinônimos: Marmaroxylon racemosum, Pithecellobium racemosum.
39 Planta tipicamente de platôs das regiões tropicais (Amazônia e Guiana), pertencente à família Leguminosae: Mimosoideae. É popularmente conhecida como angelim rajado, ingarana e ingarana da terra firme. A árvore pode atingir o dossel (25-30 m de altura) com diâmetro entre 25 a 60 cm, sem presença de sapopemas. Tronco avermelhado com manchas grandes esbranquiçadas, semelhantes a Dinizia excelsa, porém de menor porte (Ribeiro et al., 1999). A espécie é importante para a indústria, dada a dureza de sua madeira. Atualmente o angelim rajado encontra-se na lista do IBAMA de espécies protegidas, classificada como espécie vulnerável à extinção, em decorrência da exploração de sua madeira (Barcelar-Lima et al., 2006). A espécie consta na lista de plantas tropicais que são colonizadas por fungos micorrízicos arbusculares (St. John, 1980). Andira micrantha Ducke. É uma árvore de dossel de ocorrência ocasional em platôs, vertentes e campinaranas da Amazônia, pertence à família Leguminosae: Papilionoideae. É popularmente conhecida como sucupira. A árvore tem tronco circular e ritidoma marrom, com estrias verticais. Apresenta exsudação escassa, incolor, pegajosa e com forte odor de legume (Ribeiro et al., 1999). A madeira dessa espécie é bastante apreciada comercialmente. Amostragem das raízes De cada indivíduo selecionado foram coletadas amostras de raízes. O procedimento consistiu da escavação na base das plantas até encontrar a raiz principal e a partir daí segui-la até se chegar às raízes mais finas (< 2 mm) (St. John & Uhl, 1983). As raízes coletadas foram armazenadas em sacos de papel identificados e levadas para o Laboratório de Bioquímica do Solo no INPA, onde foram submetidas às análises. Colonização micorrízica A colonização por FMAs nas raízes foi estimada conforme o seguinte procedimento: as raízes coletadas foram primeiramente clarificadas e coradas pelo método de Phillips & Hayman (1970), adaptado para o uso com seringas teladas por Classen & Zasoski (1992). As raízes foram lavadas e cortadas em pedaços de aproximadamente 1 cm, dos quais 100 pedaços foram
40 selecionados ao acaso. Os segmentos radiculares foram colocados em seringas de 50 ml com pontas teladas (malha de 250 µm), submetidos a uma solução clareadora de hidróxido de potássio (KOH) 85% e levados ao banhomaria (90º C por 1h) para facilitar a penetração do reagente. Após esse processo, as raízes foram lavadas para retirada do excesso da solução de KOH. Em seguida, foram colocadas em solução clareadora de peróxido de hidrogênio alcalino (H2O2) a 30% por no máximo 20. Ainda dentro das seringas, as amostras de raízes foram novamente lavadas e cobertas com ácido clorídrico (HCl) 1% por um período de 1 h. Após a acidificação, as amostras foram cobertas com solução de azul de tripan a 0,05% em lactoglicerol acídico (na proporção de 1:10 ml respectivamente) e então retornadas ao banho-maria (90 ºC) por 1h. Após este período, as amostras permaneceram submersas em corante por mais 12 h. Ao final, 10 pedaços de raízes de cada amostra foram montadas em lâminas usando álcool polivinílico 75%. As raízes clarificadas foram avaliadas quanto à colonização micorrízica pelo método da intersecção de McGonigle et al., (1990), que consistiu no uso de um microscópio equipado com ocular milimetrada de 100 µm (com aumento de 16 vezes e uma objetiva com aumento de 10 vezes), com o qual foi observado cada pedaço de raiz em todo seu comprimento, de 100 em 100 µm. Ao final de cada 100 µm, anotou-se se nessas intersecções as raízes eram negativas ou positivas para quaisquer estruturas dos fungos micorrízicos (hifas, arbúsculos, vesículas, etc). A porcentagem do comprimento radicular colonizado total foi calculada pela seguinte equação: PCM = nº de campos positivos nº de campos positivos + nº de campos negativos x 100 Para a categorização quanto à colonização, as espécies foram classificadas em alta, média, baixa e sem colonização, quando apresentaram PCM > 50%, 49-20%, 19-1% de colonização e ausência de colonização, respectivamente (Carneiro et al.1998).
41 Amostragem do solo Com o auxílio do trado, foram coletadas amostras de solos da rizosfera de Eperua grabriflora, Zygia racemosa e Andira micrantha, em quatro pontos localizados na base de cada árvore, que consistiu de quatro sub-amostras que formaram uma única amostra composta. Estas sub-amostras foram coletadas em uma profundidade de até 10 cm, acondicionadas em sacos plásticos e levadas ao INPA para análises físicas e químicas no Laboratório Temático de Solos e Plantas. Com o auxílio de uma pá de jardim foram também coletadas nos mesmos locais, amostras com cerca de 50 g de solo para extração e contagem de esporos de FMAs no Laboratório de Bioquímica do Solo/ INPA. Densidade de esporos As amostras de solo destinadas à extração de esporos foram submetidas à secagem ao ar, sendo depois limpas para retirada das impurezas (madeira e restos vegetais). Foram pesados 10 g de solo para extração de esporos, que passaram por peneiramento úmido (Gerdemann & Nicolson, 1963) seguido pelo método de centrifugação em sacarose (Daniel & Skipper, 1982). Cada amostra de solo foi misturada em água corrente e agitada vigorosamente, seguindo-se decantação por alguns segundos, para sedimentação das partículas maiores e/ou mais densas que os esporos. A solução foi filtrada em um conjunto de peneiras de metal com abertura de 250 µm, 106 µm, 53 µm e 45 µm. O material retido foi transferido para tubos de centrífuga de 50 ml contendo solução de sacarose (50%) e submetido à centrifugação (1570 rpm por 5 ). O sobrenadante contendo os esporos foi novamente peneirado (45 µm) e lavado em água corrente, seguindo-se uma filtração em papel de filtro tipo 10, o qual foi posteriormente transferido para uma placa de Petri. Com auxílio de um microscópio estereoscópico (40x) foi efetuada a separação e contagem direta dos esporos viáveis, ou seja, aqueles cujas estruturas não estavam visivelmente deterioradas. Os esporos viáveis foram montados em lâminas com álcool polivinílico 75% (Sieverding, 1991) para futura identificação taxonômica.
42 Análises químicas e físicas do solo As amostras de solo das rizosferas das espécies selecionadas foram submetidas às análises de textura, onde foram separadas frações de argila (partículas <0,002 mm de diâmetro), silte (0,002-0,05 mm) e areia total (0,05-2 mm), usando-se o método da pipeta. A umidade foi medida por gravimetria. Avaliaram-se ainda o ph (água), o N total (extração por digestão sulfúrica, destiladores), o P disponível (extração por Mehlich 1, seguido de determinação por colorimetria espectrofotômetro ), o K disponível (extração por Mehlich 1, seguido por determinação em absorção atômica), Alumínio, Cálcio, Magnésio, Ferro, Zinco e Manganês disponíveis (extração por KCl 1 mol. L-1, seguido de determinação por absorção atômica). Todas as análises foram realizadas no Laboratório Temático de Solos e Plantas (LTSP) no INPA, segundo metodologia revisada por Embrapa (1999). Amostragem e análise química dos tecidos foliares Para análise de nutrientes nos tecidos foliares foram coletadas amostras de folhas maduras situadas em vários pontos da copa de cada indivíduo de E. grabriflora, Z. racemosa e A. micrantha, considerando folhas com boa aparência fitossanitária. Estas folhas foram armazenadas em sacos de papel previamente identificados e encaminhadas às dependências do INPA. As folhas foram lavadas em água destilada e colocadas em sacos de papel para serem secas em estufa com circulação forçada de ar a 60 ºC por 72 h. Em seguida, foram trituradas em moinho do tipo Wiley, passadas em peneira de malha de 1,0 mm (20 mesh) e armazenadas em frascos de vidro. Os tecidos foliares foram analisados quimicamente quanto aos teores de macro e micronutrientes (nitrogênio, fósforo, potássio, cálcio, magnésio, ferro, zinco e manganês), usando a metodologia descrita em Embrapa (1999). Análise dos dados A colonização micorrízica e a densidade de esporos presentes na rizosfera, nas três espécies de Leguminosae (Eperua grabriflora, Zygia racemosa e Andira micrantha) foram comparadas entre si por análise de variância (ANOVA) e as suas médias submetidas ao teste de Tukey a 95% de significância.
43 As propriedades físicas e químicas do solo (macro- e micronutrientes, ph, umidade e textura do solo) e macro- e micronutrientes presentes nos tecidos foliares foram ordenados por Análise de Componentes Principais (PCA), que resultou em três eixos para cada espécie analisada. Estes eixos foram ajustados usando-se o teste de Bonferroni e testados por correlação de Pearson. Análises de regressão linear foram realizadas entre as taxas de colonização micorrízica das três espécies e as propriedades físicas e químicas do solo, primeiramente com os três eixos de PCA ordenados destas variáveis e em seguida com essas variáveis individualmente. Para avaliar a contribuição das micorrizas na alocação de nutrientes nos tecidos foliares, também foram realizadas análises de regressão linear entre as taxas de colonização e os macro- e micronutrientes das folhas, primeiramente quando ordenados nos eixos de PCA e em seguida com os nutrientes individualmente. Análises de regressão múltipla também foram utilizadas pra examinar a relação entre as densidades de esporos encontrados na rizosfera de cada espécie de Leguminosae e as propriedades químicas e físicas do solo (nutrientes, ph, umidade e textura do solo), ora ordenados em eixos de PCA, ora individualmente. Por fim, a relação entre as altitudes das parcelas e os fatores taxas de colonização micorrízica e densidade de esporos também foi avaliada por meio de análise de regressão linear. Todas as análises estatísticas foram realizadas no programa SYSTAT 10.0 for Windows (Wilkinson, 1998).
44 RESULTADOS Propriedades do solo A umidade do solo ao longo do gradiente topográfico apresentou grande variação entre as parcelas (Figura 6 A). Quanto à textura do solo, no geral as parcelas que se encontravam em baixas altitudes apresentaram maiores teores de areia (Figura 6 B), onde foi encontrada a maioria dos indivíduos de E. glabriflora e A. micrantha. Nesses locais, os solos apresentaram médias de 69 e 61%, respectivamente, ao passo que os solos onde foram encontradas as plantas de Z. racemosa apresentaram maiores teores de silte (65,8%). A topografia de todas as parcelas foi fortemente correlacionada com as variáveis de textura do solo (% de argila, areia, silte). Não houve variações significativas do ph do solo nas parcelas (3,9 a 5,6). Classificando-os como solos ácidos (Figura 6 C). Em geral, os solos das parcelas onde os indivíduos de E. glabriflora foram amostrados apresentaram as maiores concentrações de macronutrientes e nas parcelas onde foram amostradas as plantas de Z. racemosa, foram encontradas as maiores concentrações de micronutrientes. Contudo, as concentrações de P e Fe2+ nas parcelas amostradas para esta espécie foram correlacionadas negativamente com a altitude (r = -0, 668, p < 0,001; r = -0, 516, p < 0, 001, respectivamente). Indivíduos de A. micrantha foram encontrados nas parcelas com solos mais pobres em macro- e micronutrientes. O Apêndice 1 mostra as propriedades físicas e químicas do solo das rizosferas das plantas de E. glabriflora, Z. racemosa e A. micrantha amostradas. Análise dos Componentes Principais (PCA) para variáveis de solo e nutrientes nas folhas A análise de ordenação revelou três eixos principais das variáveis do solo para cada espécie estudada (Tabela 1). Em E. glabriflora o Eixo 1, que explicou 24% do total da variação, descreve principalmente as variáveis K+, Ca2+, Mg2+, Zn2+ e Mn2+. O Eixo 2 explicou 19% da variação, e foi influenciado pelas variáveis silte, argila, P e Fe2+ e o Eixo 3 que explicou 12% da variação, descreve principalmente as variáveis % de umidade, ph, P, K+ e Al3+.

45 A B C D E F Figura 6. Distribuição de algumas variáveis físicas e químicas do solo ao longo de um gradiente de altitude de 30 parcelas na RFAD.
46
47 Tabela 1. Correlação de Pearson entre 13 variáveis do solo (de 0 10cm) e 8 nutrientes foliares e três eixos de ordenação produzidos da Análise de Componentes Principais (PCA). Variáveis do solo Eperua glabriflora Zygia racemosa Andira micrantha Eixo 1 Eixo 2 Eixo 3 Eixo 1 Eixo 2 Eixo 3 Eixo 1 Eixo 2 Eixo 3 0,305-0,076 0,553** 0,505* -0,645** 0,118 0,227-0,413 0,739** 0,132-0,890** 0,208-0,890** 0,143-0,157 0,799** -0,506 0,024 Argila (%) 0,259-0,877** 0,260-0,774** 0,019 0,153 0,711* -0,566 0,154 ph (H20) 0,151-0,081 0,443** -0,170-0,367 0,146 0,551-0,129-0,650 N 0,249 0,326 0,235 0,032 0,426 0,133-0,098 0,375 0,138 P -0,164 0,639** 0,523** 0,736** -0,561** 0,147-0,652 0,005 0,666 K 0,563** 0,004 0,476** -0,298 0,018 0,769** 0,084-0,584 0,542 0,889** 0,224-0,080 0,167 0,582** 0,211 0,568 0,674 0,221 0,820** 0,237-0,078 0,273 0,670** 0,076 0,638 0,655 0,148 0,088-0,092-0,441** -0,075-0,145-0,425 0,307 0,274 0,271 Umidade (%) Silte (%) + Ca 2+ Mg Al Fe 2+ 0,315-0,495** -0, ,123 0,780** 0,715* -0,375-0,106 Zn2+ 0,767** 0,110-0,363 0,677** 0,399-0,089 0,644 0,213 0,095 Mn2+ 0,649** 0,135-0,064 0,459 0,452 0,139 0,515 0,594 0,342 Altitude (m) 0,100-0,871** 0,160-0,888** 0,095-0,048 0,750** -0,538 0,136 Variação explicada (%) Nutrientes foliares N 0,044 0,112 0,715** 0,383** 0,403-0,550** -0,516 0,417 0,316 P 0,308 0,771** -0,342 0,780** 0,015 0,100-0,889** 0,239-0,008 K 0,464** 0,639** -0,454** 0,657** 0,103 0,193-0,647** 0,531 0,424 0,831** -0,315 0,182 0,597** -0,588** 0,016 0,470 0,655* 0,046 0,903** -0,111 0,121 0,579** -0,465** -0,270 0,518 0,675* 0,296 0,046 0,555** 0,521** -0,338-0,403 0,682** 0,723** 0,268 0,206-0,062 0,642** 0,407* 0,638** 0,167 0,500* -0,491 0,427-0,578 0,809** -0,201 0,031-0,182-0,791** -0,348 0,241 0,537-0,733** Ca 2+ Mg 2+ Fe 2+ Zn 2+ Mn 2+ Variação explicada (%) Obs: Teste de Bonferroni: ** significante a 0,01; * significante a 0,05
48 Tanto para E. glabriflora como para Z. racemosa o Eixo 1 explicou 25% da variação total. Para E. glabriflora, esse eixo descreveu principalmente as variáveis nutricionais K+, Ca2+, Mg2+, Zn2+ e Mn2+. Para Z. racemosa o Eixo 1 descreveu principalmente as variáveis físicas de umidade, silte e argila e as variáveis nutricionais de P e Zn2+ das parcelas. O Eixo 2 explicou 19 e 17% para E. glabriflora e Z. racemosa, respectivamente, sendo que para Z. racemosa esse eixo refletiu principalmente as variáveis de umidade, P, Ca2+, Mg2+ e para E. glabriflora esse eixo refletiu mais as variáveis argila, silte, P e Fe2+. O Eixo 3 explicou 12% da variação para as duas espécies e descreveu somente as varáveis K+ e Fe2+ em Z. racemosa e em E. glabriflora as variáveis de umidade, ph, P, K+ e Al3+. Para A. micrantha o Eixo 1 explicou mais de 30% da variação e foi relacionado principalmente com os teores de silte, argila e Fe2+. O Eixo 2 não estava relacionado significativamente com nenhuma variável, porém explicou 21% da variação. O Eixo 3 explicou 16% da variação e descreveu fortemente a variável de umidade do solo nas parcelas onde estavam as plantas de A. micrantha. No mínimo um eixo de cada uma das três espécies foi correlacionado com os valores de altitudes nas parcelas. Em E. glabriflora o Eixo 2 foi negativamente correlacionado com altitude (r =-0,871; p < 0,001). Em Z. racemosa o Eixo 1, que refletiu principalmente as variáveis físicas do solo e os nutrientes P e Zn2+, também foi inversamente correlacionado com a altitude das parcelas (r = ; p < 0,001) e em A. micrantha o Eixo 1 também foi correlacionado com a topografia (r = 0,750; p = 0.001). Também foram realizadas análises de ordenação para as variáveis nutricionais das folhas das três espécies amostradas nesse estudo (Tabela 1). Em E. glabriflora os três eixos principais explicaram juntos mais de 72% da variação. O Eixo 1 refletiu principalmente as variáveis nutricionais K +, Ca2+, Mg2+, Mn2+ e explicou 31% dessa variação, enquanto que o Eixo 2 explicou 24% da variação e refletiu principalmente as variáveis P, K+, Fe2+, Zn e por último o Eixo 3 refletiu os nutrientes N, K+, Fe2+ e Zn2+ e explicou 17% da variação. O PCA de Z. racemosa explicou quase 90% da variação, com o Eixo 1, refletindo as variáveis de N, P, K+, Ca2+, Mg2+, Zn2+. O Eixo 2 refletiu principalmente Ca2+, Mg2+, e Mn2+ e explicou 19% da variação. Por sua vez, o Eixo 3 explicou 6% da variação e refletiu somente os nutrientes N, Fe2+, Zn2+.
49 O Eixo 1 de A. micrantha explicou 35% da variação e mostrou forte correlação com as variáveis P, K+ e Fe2+. O Eixo 2 explicou 24% da variação e foi mais fortemente relacionado com as variáveis Ca2+ e Mg2+, enquanto que o Eixo 3 só foi relacionado com Mn2+ e explicou apenas 16% da variação total. Colonização micorrízica em E. glabriflora, Z. racemosa e A. micrantha As médias das porcentagens de colonização do comprimento de raízes das espécies de Leguminosae estudadas foram similares entre E. glabriflora e A. micrantha, com 63 e 52%, respectivamente, sendo classificadas como espécies altamente colonizadas. Por sua vez, Z. racemosa apresentou baixa colonização, com apenas 9% do comprimento das suas raízes colonizadas. Em E. glabriflora a colonização das raízes por FMA obteve ampla variação de 45,5 a 75,5%, porém não apresentou diferença significativa (ANOVA, F = 1,581; p = 0,111) entre si; assim como em Z. racemosa (ANOVA, F = 1,307; p = 0,259) onde a colonização micorrízica variou de 2,75 a 13,4% e Porcentagem de Colonização Micorrízica (%) em A. micrantha (ANOVA, F = 0,752; p = 0,596) de 42 a 61% (Figura 7) Eperua glabriflora Andira micrantha Zygia racemosa Espécies Figura 7. Porcentagem média de colonização micorrízica em seguimentos de raízes de indivíduos de Eperua. glabriflora, Zygia. racemosa e Andira micrantha. A colonização micorrízica de E. glabriflora, Z. racemosa e A. micrantha não foi relacionada com a altitude das parcelas e nem com as características do solo quando ordenadas nos três eixos de PCA. A ausência de relação
50 também foi observada entre a colonização micorrízica e os três eixos de PCA ordenados das variáveis de nutrientes foliares (Tabela 2). Tabela 2. Regressão linear entre as porcentagens de colonização micorrízica - PCM nas raízes de E. glabriflora, Z. racemosa e A. micrantha e as características do solo (eixos de PCA). Regressão Linear PCM x Altitude PCM x PCA 1 solo PCM x PCA 2 solo PCM x PCA 3 solo PCM x PCA 1 foliar PCM x PCA 2 foliar PCM x PCA 3 foliar Eperua glabriflora 0,001 0,002 0,019 0,024 0,011 0,017 0,005 Valores de r2 Zygia racemosa Andira micrantha 0,000 0,002 0,048 0,016 0,001 0,074 0,006 0,068 0,070 0,019 0,116 0,051 0,005 0,000 A análise de regressão linear entre as porcentagens de colonização micorrízica nas raízes de E. glabriflora e as variáveis do solo (macro- e micronutrintes, textura, umidade e ph) analisadas separadamente também não mostraram ter qualquer relação. Para as outras duas espécies dois nutrientes do solo influenciaram a colonização micorrízica nas suas raízes. Em Z. racemosa houve relação significativa com as concentrações de Fe2+, enquanto que em A. micrantha o K+ foi o nutriente que influenciou significativamente as porcentagens de colonização (Figura 8). y = x R2= 0.083* A y = 0.519x R2= 0.181* B Figura 8. Relação entre a colonização micorrízica (%) (A) nas raízes de Z. racemosa e as concentrações de Fe2+ (mg.kg-1) do solo; (B) nas raízes de A. micrantha e as concentrações de K+ (mg.kg-1).
51 Os nutrientes presentes nos tecidos foliares coletados das três espécies de Leguminosae (Tabela 3) também não foram relacionados com as porcentagens de colonização micorrízica das raízes das três espécies como mostraram os resultados das regressões lineares realizados com os valores da colonização micorrízica (%) e os nutrientes presentes nas folhas testados individualmente. Tabela 3. Concentrações de macro- e micronutrientes presentes em folhas de E. glabriflora, Z. racemosa e A. micrantha. Os números são médias e desvios padrão (entre parênteses). Espécies E. glabriflora (n = 69) Z. racemosa (n = 49) A. micrantha (n = 23) N Total (%) 1,34 Macronutrientes K+ Ca2+ -1 g.kg 0,77 3,87 1,24 P Mg2+ 0,87 Micronutrientes Zn+ Mn2+ -1 mg.kg 26,28 12,08 129,21 Fe2+ (±0,21) (±0,15) (±1,46) (±0,64) (±0,25) (±8,15) (±4,87) 2,56 0,71 1,85 0,93 0,60 73,43 9,02 (±72,95) 50,54 (±0,33) (±0,13) (±0,78) (±0,34) (±0,16) (±33,92) (±2,06) (±20,96) 1,74 0,84 1,83 0,86 0,57 47,76 14,84 62,51 (±0,26) (±0,29) (±1,15) (±0,59) (±0,23) (±25,69) (±6,29) (±46,11) Densidade de esporos presentes na rizosfera de E. globriflora, Z. racemosa e A. micrantha A densidade de esporos presentes nos solos coletados na rizosfera das três espécies de Leguminosae foi muito variada. Em E. glabriflora, a amplitude de variação da densidade de esporos foi de esporos/ 10 g de solo; em Z. racemosa, a densidade de esporos variou de esporos/ 10 g de solo e na rizosfera de A. micrantha, a amplitude de variação da densidade de esporos foi de de esporos/ 10 g de solo (Figura 9). Apenas em E. glabriflora houve variação intra-específica significativa (ANOVA, F = 2,022; p = 0,031).
52 Densidades Médias de Esporos/ 10.g -1 de solo Eperua glabriflora Andira micrantha Zygia racemosa Espécies Figura 9. Densidade média de esporos em 10 g de solo coletado na rizosfera de Eperua glabriflora, Zygia racemosa e Andira micrantha. As análises de regressão linear entre as densidades de esporos na rizosfera de E. glabriflora e A. micratha e os eixos do PCA não mostraram relação significativa. Porém, em Z. racemosa foi obtida relação entre o Eixo 2, que refletiu principalmente as variáveis edáficas de umidade, P, Ca2+ e Mg2+ e a densidade de esporos na rizosfera dessa espécie (F = 4,612; p = 0,037). Tabela 4. Regressão linear (r2) da densidade de esporos (DE) em E. glabriflora, Z. racemosa e A. micrantha e os três eixos ordenados de PCA das variáveis do solo. Regressão linear DE x Altitude DE x Eixo 1 solo DE x Eixo 2 solo DE x Eixo 3 solo Eperua glabriflora 0,005 0,002 0,000 0,010 Valores de r2 Zygia racemosa 0,031 0,009 0,165 0,175 Andira micrantha 0,315 0,100 0,144 0,002 Também houve relação entre a densidade de esporos e três macronutrientes pelos testes de regressão linear das densidades de esporos com P total, Ca2+ e Mg2+, individualmente (Figura 10A, B, C).
53 y = x R2 = * A y = x R2= 0.099* y = x R2 = * B C Figura 10. Relações entre densidade de esporos presentes na rizosfera de Z. racemosa e os macronutrientes (mg. Kg-1) no solo, Fósforo (A), Cálcio (B) e Magnésio (C). As análises de regressão entre as propriedades físicas e químicas do solo das parcelas e as densidades de esporos presente nas rizosferas mostraram relações para as três espécies. Em E. glabriflora houve relação significativa entre a densidade de esporos e o ph do solo (Figura 11). Em A. micrantha a densidade de esporos foi relacionada com fatores físicos do solo (Figura 12A, B), como a altitude, % areia, %silte e % argila.
54 y = x R2= * Figura 11. Relação entre densidade de esporos presentes na rizosfera de E. glabriflora e o ph do solo. y = 0.778x R2 = ** A y = x R2 = ** B Figura 12. Relações entre densidade de esporos presentes na rizosfera de A. micrantha e a altitude (gráfico A) e % de argila (gráfico B). A taxonomia das espécies de fungos micorrízicos, feita a partir da montagem dos esporos, não foi realizada nesse estudo por falta de tempo hábil. Porém as lâminas são permanentes e estão armazenadas para futura identificação.
55 DISCUSSÃO Colonização de MA nas raízes de Eperua glabriflora, Zygia racemosa e Andira micrantha Foi encontrada uma grande amplitude de variação entre a colonização MA em E. glabriflora (45-75%), Z. racemosa (2-13%) e A. micrantha (42-61%). Em E. glabriflora e A. micrantha os valores observados estão dentro da faixa de média a alta colonização, fato também observado em outras espécies de leguminosas (Bonetti et al. 1984, Sieverding, 1991, Honrubia et al., 1992; Grove & Le Tacon, 1993). A baixa colonização micorrízica em Z. racemosa, entretanto contrastam com as obtidas por Bonetti et al. (1984) em seu estudo dessa espécie em plantios florestais na Amazônia, onde obtiveram alta colonização (75 86%), sugerindo que a associação micorrízica nessa espécie varia conforme o ambiente. Frioni et al. (1999) analisaram diversas espécies de árvores e arbustos das três subfamílias de Leguminosae provenientes de diferentes regiões do Uruguai e também obtiveram colonização com alta amplitude de variação para a subfamília Caesalpinioideae (6-74%), à qual pertence E. glabriflora. Os percentuais obtidos por eles para as subfamílias Papilionoideae (62-78%) e Mimosoideae (18-69%) foram maiores do que os encontrados para A. micrantha (Papilionoideae) e Z. racemosa (Mimosoideae) neste estudo. Em casa de vegetação, diferentes graus de colonização para a família Leguminosae foram obtidos por Carneiro et al. (1998) e várias espécies (cerca de 15%, n = 38) dessa família foram consideradas como não-micotróficas. Para os autores, baixas porcentagens de colonização em leguminosas podem ser intrínsecas, visto que análises de outros fatores não apresentaram nenhuma relação com esses índices. Esse argumento também foi usado por Siqueira & Saggin-Júnior (2001), sugerindo que as espécies de plantas e fungos envolvidos nessa associação mutualística se expressam em diferentes graus de colonização nas plantas hospedeiras de comunidades similares, ou seja, não há especificidade de hospedeiro e nem padrão de colonização. Em cultivo no campo, Zangaro et al. (2003) também encontraram baixas porcentagens de colonização em diversas espécies de Leguminosae da bacia do rio Tibagi, Paraná, Brasil. Por exemplo, 1% para Pterogyne nitens (Caesalpinioideae), 7,2% para Centrolobium tomentosum (Papilionoideae),
56 3,3% para Machaerium Lonchocarpus muehlbergianus stipitatum (Papilionoideae) (Papilionoideae), 3,1% para e 4,2% para Inga striata (Mimosoideae). Tanto esse estudo (Zangaro et al., 2003) como de Siqueira et al. (1998), mostraram um padrão de resposta das espécies lenhosas de diferentes grupos sucessionais, isto é, a colonização foi muito maior em espécies pioneiras e secundárias iniciais do que em espécies secundárias tardias e em clímax. As três espécies deste estudo pertencem ao grupo sucessional de espécies clímax. A espécie pertencente à subfamília Mimosoideae, Anadenanthera colubrina, testada em diferentes níveis de fertilidade em casa de vegetação, também apresentou baixa colonização micorrízica (0-14%), não correlacionadas com a fertilidade do solo do local (Patreze & Cordeiro, 2004). Segundo os autores, estas porcentagens de colonização poderiam estar mais associadas às especificidades da planta hospedeira e do fungo do que com as condições nutricionais. As micorrizas são de ocorrência generalizada nas plantas vasculares (Brundrett, 1991), porém existem espécies que são pouco suscetíveis à colonização ou não dependem da simbiose para o crescimento. Em ambiente natural de cerrado, os baixos índices de colonização de espécies nativas desse bioma têm sido explicados pela baixa incidência de esporos de FMAs encontrados no solo, em média 8 esporos/50 g de solo (Carneiro et al., 1998). Este, porém, não foi o caso para a espécie Z. racemosa, cuja densidade de esporos foi relativamente alta, em média 51 esporos/10 g de solo extraídos de sua rizosfera. Em uma floresta primária da Guiana Francesa, foram encontradas porcentagens de colonização micorrízicas de 60% nas raízes de Eperua falcata (Martin et al., 2001). Essa porcentagem é similar à encontrada para uma das espécies do presente estudo do mesmo gênero, a Eperua glabriflora (45 75%). A topografia florestal está claramente associada a diversas variáveis ambientais, principalmente àquelas relacionadas ao solo e são determinantes para a estruturação vegetal nestes ambientes, como descrito por inúmeros estudos realizados em várias escalas (Laurance et al., 1999; Clark & Clark, 2000; Takyu et al., 2003; Costa et al. 2005; Daws et al., 2005; Castilho et al, 2006). Na RFAD a porcentagem de argila no solo está relacionada com a topografia (Chauvel et al., 1987; Mertens, 2004) e com a distribuição espacial
57 de plantas (herbáceas Costa et al., 2005 e Costa, 2006; espécies de subbosque - Kinupp & Magnusson, 2005). Por isso, esperava-se encontrar fortes relações entre a colonização micorrízica arbuscular nas raízes das três espécies de Leguminosae analisadas no presente estudo e as variáveis edáficas. Espécies vegetais arbóreas distribuídas em um gradiente topográfico de uma região do Arizona também não foram correlacionadas com MA (Fisher & Fulé, 2004). O estudo apenas testou o efeito da topografia, não considerando todas as variáveis envolvidas na variação da altitude como o microclima, a cobertura do dossel, a luminosidade, a área basal arbórea, os nutrientes do solo, a umidade do solo, a ciclagem de nutrientes da liteira no solo e espécies do fungo MA (Benjamin et al., 1989; Johnson et al., 1991; Anderson et al., 1994; Schultz et al., 2001; Wood et al., 2006). As mudanças causadas pela elevação em um gradiente acabam por mudar a umidade do solo, temperatura, inclinação, a estrutura florestal, vegetação e nutrientes do solo (Austin, 1999). A ausência de respostas significativas da colonização micorrízica nas raízes de E. glabriflora com as propriedades físicas e químicas do solo no presente estudo foi inesperada, pois a simbiose entre os organismos envolvidos é baseada na troca de nutrientes entre ambos. Estes resultados contradizem os de Frioni et al. (1999), que concluíram que a relação entre árvores de Leguminosae e o fungo micorrízico varia de espécie para espécie e com as condições ambientais e, acima de tudo, com a fertilidade do solo. Sieverding (1991) também sugeriu que o grau de colonização micorrízica é dependente das condições do solo, embora outras fontes de variação inclusive da própria planta hospedeira e das espécies de FMA presentes no local, tenham sido consideradas. Embora inesperada, não é a primeira vez que essa ausência de efeito dos elementos minerais, incluindo o fósforo, sobre a colonização micorrízica foi observada. Eperua falcata cultivada em ambientes controlados na Guiana Francesa (Grandcourt et al., 2004) foi considerada micotrófica facultativa na fase de plântula, por não mostrar um crescimento dependente da colonização micorrízica e nem responder à adição de fósforo. Aparentemente, essa ausência de dependência da colonização para absorção de fósforo no gênero de Eperua pode ser extensiva a E. glabriflora quando adultas, conforme evidenciado no presente estudo.
58 Os resultados no presente estudo e em outros sugerem que na associação planta hospedeira x FMA, o crescimento de ambos os organismos envolvidos na simbiose é influenciada por outros fatores relacionados a planta hospedeira, mas não necessariamente está relacionado com a fertilidade do solo (Bethlendfalvay et al., 1985) ou com a porcentagem de colonização (Vierheilig & Ocampo, 1991). A relação negativa das concentrações de Fe2+ e a colonização micorrízica em Z. racemosa e a relação positiva das concentrações de K+ e as porcentagens de colonização micorrízica de A. micrantha, mostraram que pelo menos para essas duas espécies de Leguminosae as conclusões de Frioni et al. (1999), Sieverding (1991) e outros autores, de que alguns elementos minerais do solo influenciam a ocorrência dessa associação nos hospedeiros, estejam ocorrendo nesse estudo. Embora estas relações possam ter sido casuais para ambos, os elementos e a espécie, as árvores de Z. racemosa continham as maiores concentrações de ferro que as outras duas espécies. Caris et al. (1998) sugeriram que o Fe2+ é um importante nutriente que pode ser absorvido pelas hifas de FMA e transferido para plantas hospedeiras. Isso também foi observado em um experimento com um cultivar de Zea mays susceptível à deficiência de Fe2+, quando a colonização micorrízica aumentou significativamente as concentrações de ferro na planta (Clark & Zeto, 1996). A relação entre espécies de Leguminosae e os microrganismos do solo é um fator importante a ser considerado em estudos do comportamento dessa família nos variados ambientes. Além dos fungos micorrízicos que favorecem o crescimento plantas com a absorção de nutrientes (Abbott & Gazey, 1994), as leguminosas também estabelecem associação com rizóbios que favorecem a nodulação e absorção principalmente de nitrogênio, entre outros benefícios (Silva et al., 1994). As leguminosae acabam sendo mais exigentes por fósforo, em parte devido a sua capacidade de fixar N2.. A dupla associação simbiótica com rizóbio e fungo micorrízico deixam as leguminosas mais eficientes na obtenção de nutrientes minerais e água e conseqüentemente elas podem competir com outras plantas (Frioni et al., 1999). Neste estudo não foi possível avaliar até que ponto a associação com bactérias fixadoras de N2 podem ter mascarado a avaliação das associações das plantas com FMA.
59 A ausência do efeito benéfico da colonização micorrízica no aumento das concentrações de macro- e micronutrientes nos tecidos foliares de E. glabriflora, Z. racemosa e A. micrantha, mostrou que o acúmulo de nutrientes é aparentemente independente da colonização micorrízica. Esses resultados contrastam com diversos estudos que demonstraram correlações entre essas variáveis em condições naturais (Colozzi-Filho & Siqueira, 1986; Oliveira et al. 1999; Oliveira et al. 2003). Entretanto, outras observações como a de Oliveira et al. (1999), por exemplo, não encontraram relação entre a colonização micorrízica e os nutrientes foliares em Dipterix odorata, Eremoluma williamii, Scleronema micranthum e Cedrelinga catenaeformis em condições naturais. Respostas simililares também foram obtidas por Carneiro et al. (1998), Siqueira et al. (1998) e Cuenca et al. (2003), quando as concentrações de macro- e micronutrientes dos tecidos foliares não apresentaram variação em resposta a colonização micorrízica. Densidade de esporos de FMA nas rizosferas de E. glabriflora, Z. racemosa e A. micrantha Somente a presença do esporo na rizosfera não é um indicador de eficiência da associação deste com a planta hospedeira, pois os esporos são células de resistência produzidas em resposta a algum estresse ambiental. Bever et al. (1996) e Bever (2002) demonstraram que cada esporo de FMA pode ser multiplicado diferentemente nas diversas plantas hospedeiras e essas associações podem ser tanto positivas como negativas, ocorrendo um feedback entre uma planta hospedeira específica e seus organismos endofíticos. Para que o processo de esporulação seja eficiente é necessário que haja grande quantidade de compostos orgânicos, principalmente lipídios, o que ocasiona acentuado dreno destes compostos para o fungo (Peng et al., 1993). As variações intra-específicas na densidade de esporos na rizosfera de E. glabriflora, Z. racemosa e A. micrantha foram maiores do que as interespecíficas mascarando quaisquer possíveis relações significativas com as taxas de colonização. Resultados similares também foram obtidos por Martin et al. (2001) em que o grau de colonização micorrízica nas raízes de Dicorynia
60 guianensis e Eperua falcata não teve relação com a densidade de esporos nas rizosferas dessas espécies. As densidades de esporos encontrados nas rizosferas de E. glabriflora diferiram intra-especificamente e foram relacionadas positivamente com ph do solo nas parcelas. Outros estudos (Mendonza et al., 2002; Mohammad et al., 2003; Escudero & Mendonza, 2005, Isobe et al., 2007) também encontraram relação positiva entre ph do solo e as densidades de esporos. Robson & Abbott (1989) reportam que algumas espécies de FMA são restritos a solos ácidos e alcalinos ou ocorrem em ambos. Não houve variação nas densidades de esporos nas rizosferas de Z. racemosa e nem na de A. micrantha, mas ambas foram relacionadas com a variação das propriedades físicas e químicas do solo. Em Z. racemosa as densidades de esporos foram influenciadas negativamente com as concentrações de P e positivamente pelas concentrações de Ca2+ e Mg2+. Mohammad et al. (2003) reportaram que a quantidade de P no solo afetou negativamente a produção, a sobrevivência e a germinação de esporos de FMA. Outros autores também obtiveram correlações similares entre a quantidade de nutrientes, principalmente o P e a densidade de esporos de FMA no solo (Mendonza et al., 2002; Pande & Tarafdar, 2004). Considerando que os esporos são células de resistência, em antecipação a estresse ambiental seria esperado que sua produção aumentasse quando os nutrientes do solo se tornassem escassos. As densidades de esporos presentes na rizosfera de A. micrantha foram influenciadas pela altitude das parcelas e pelas variáveis de textura do solo como % de areia, de silte e de argila. O tipo de solo também afetou a densidade de esporos em outros estudos, como por exemplo, Lekberg et al. (2007) obtiveram respostas da influência da textura do solo no local de estudo sobre a distribuição de diferentes espécies. Os resultados encontrados em E. glabriflora, Z. racemosa e A. micrantha para as densidades de esporos sugerem que possivelmente os FMA que estão colonizando essas espécies sejam de diferentes famílias taxonômicas visto que respondem diferentemente as variáveis testadas. Em condições naturais a composição das comunidades de fungos micorrízicos difere grandemente entre tipos de solo e seus constituintes de fertilidade, como relatado por Lekberg et al. (2007), quando avaliaram o comportamento de espécies de duas famílias de
61 fungos MA em solos de diferentes texturas e quantidades de nutrientes. Infelizmente, no presente estudo a taxonomia das espécies de FMA ainda não foi realizada para avaliar essa possível relação. Em uma floresta tropical no sudoeste da China também foram encontradas evidências de que a densidade de esporos presentes na rizosfera de 112 espécies está estreitamente relacionada com o solo e os seus complexos componentes e com os grupos de FMA presentes no local (Zhao et al., 2001). Em um estudo da distribuição espacial em pequena escala de esporos de FMA em uma vegetação na zona costeira de Portugal foi obtido alto grau de variabilidade na densidade de esporos e uma alta dependência da distribuição espacial e do tipo de vegetação nestas áreas, sugerindo que a distribuição de esporos de MA nesse caso é dependente da composição e distribuição das plantas (Carvalho et al., 2003). Em contraste, não foi encontrada nenhuma relação entre a densidade de esporos e uma planta hospedeira em um estudo de distribuição em pequena escala (Friese & Koske, 1991). Os resultados neste estudo indicam que uma variação topográfica de apenas 70 metros na altitude não induziu a estresses ambientais suficientemente drásticos para que houvesse uma esporulação marcante e diferenciada entre os indivíduos da família Leguminosae estudados. Tanto para as análises das PCM como para densidade de esporos presentes nas três espécies de Leguminosae, a identificação taxonômica dos FMA será fundamental para responder algumas importantes questões relacionadas à composição da comunidade de FMA e sua distribuição no gradiente florestal estudado. As Tabelas 5, 6 e 7 sumarizam os principais resultados dos estudos sobre micorrizas comentados no presente estudo.
62 Tabela 5. PCM e densidade de esporos (DE) de FMA associados a várias espécies vegetais na Amazônia brasileira. EspéciesAlvo Tipo de floresta FMA Local PCM DE (10 g solo) Observações Referência E. glabriflora, Z. racemosa e A. micrantha Floresta tropical Amazonas, Brasil Altas e baixas esporos Baixa influência das propriedades do solo e ausência de relação com nutrientes das folhas. Indivíduos de Sapotaceae, Burseraceae e Lecythidaceae Floresta tropical Amazonas, Brasil Médias esporos. Sem relação entre as famílias e MA. Cultivares de banana Floresta tropical Amazonas Brasil Altas e Médias C. catenaeformis, D. odorata e outras espécies florestais Floresta tropical Amazonas, Brasil Alta e Médias - 31 espécies da Família Lecythidaceae Floresta tropical Amazonas, Brasil Baixa ou Ausente - C. catenaeformis, D. odorata, D. potiophylla, C. racemosa, P. racemosum, V. palidior, E. bijuga. Floresta tropical Amazonas Brasil Altas e Médias 1,6 27,2 esporos Colonizações micorrízicas não apresentaram relação com nº de esporos. Bonetti et al., 1984 Várias espécies florestais Floresta tropical Amazonas, Pará e Bahia Brasil - Todas as espécies listadas eram micorrizadas, porém relação com nutrientes solo e planta não testada St. John, 1980 (-) Variável não avaliada - - A associação micorrízica favoreceu a absorção de nutrientes. Todas as espécies fortemente relacionadas com nutrientes foliares Ausência total de fungos nas raízes ou presença de poucas estruturas fúngicas. Este estudo Ropero, 2007 Oliveira et al., 2003 Oliveira et al Moreira et al. 1997
63 Tabela 6. PCM e densidade de esporos (DE) de FMA associados a várias espécies vegetais no Brasil. Espécies- Tipo de Alvo floresta FMA Local Alta e Baixa - Absorção de P não favoreceu o crescimento das plântulas. Patreze & Cordeiro, Acentuado micotrofismo relacionado com baixa fertilidade do solo Zangaro et al Floresta tropical Espécies de diferentes grupos sucessionais Floresta tropical Paraná, Brasil Floresta tropical Minas Gerais, Brasil Floresta tropical São Paulo Brasil - 8,5 51,1 esporos Floresta tropical semidecídua Minas Gerais Brasil Alta, Médias e Baixas 7,2 20 esporos 29 espécies lenhosas (12 leguminosas) C. urucurana, I. striata e G. americana 35 espécies de Leguminosae (-) Variável não avaliada Referência DE (10g solo) A. colubrina, M. bimucronata, P. rigida (Leguminosae Mimosoideae) São Paulo Brasil Observações PCM Altas em pioneiras e baixas em clímax Alta em 60% das espécies Ampla variação Eficiência da absorção de P foi afetada pelas MA e níveis de P. O número de esporos tendeu a aumentar com o estádio sucessional. Ocorrência generalizada de MA nas espécies arbóreas tropicais e relação com nutrientes do solo e da planta não testada. Siqueira & Saggin-Júnior, 2001 Carrenho et al., 2001 Carneiro et al. 1998
64 Tabela 7. PCM e densidade de esporos (DE) de FMA associados a várias espécies vegetais em diferentes localidades. Espécies- Tipo de Alvo floresta FMA Local PCM DE (10g solo) Observações Referência Indivíduos de Sapotaceae, Burseraceae e Lecythidaceae Floresta tropical Colômbia Médias esporos Sem relação entre as famílias e MA. Ropero, 2007 Indivíduos de Sapotaceae, Burseraceae e Lecythidaceae Floresta tropical Peru Médias esporos Sem relação entre as famílias e MA. Ropero, 2007 Dicorynia guianensis e Eperua falcata Floresta tropical Guiana Francesa Altas Dicorynia guianensis e Eperua falcata Floresta tropical Guiana Francesa Floresta tropical 112 espécies 23 espécies de árvores e arbustos de leguminosae Vários locais (-) Variável não avaliada - Presença de MA relacionado positivamente com o P presente nas folhas. Grandcourt et al Altas esporos Presença acentuada de FMA em ambas espécies e em suas rizosferas. Martin et al., 2001 China 56% das espécies altamente micorrizadas 5,5 190,8 esporos A distribuição dos esporos de FMA está relacionado positivamente com a fertilidade do solo. Zhao et al., 2001 Uruguai Altas, Médias e Baixas esporos A relação planta X MA varia com a espécie, as condições ambientais e a fertilidade do solo. Frioni et al., 1999
65 CONCLUSÕES As espécies E. glabriflora, Z. racemosa e A. micrantha responderam diferentemente à colonização micorrízica. Altas porcentagens de colonização micorrízica foram obtidas para E. glabriflora (63%) e para A. micrantha (52%), enquanto que para Z. racemosa as porcentagens de colonização micorrízica foram sempre baixas (9%). A ausência de especificidade da planta hospedeira parece gerar graus diferenciados de colonização micorrízica mesmo quando se trata de uma mesma espécie e/ou espécies que pertencem a uma mesma família taxonômica. Os resultados deste estudo invalidam a hipótese 1 de que haveria relação entre as porcentagens de colonização micorrízica nas raízes de E. glabriflora, Z. racemosa e A. micrantha e a altitude nos 25 km2 da floresta estudada. A ausência dessas relações indica que embora as plantas hospedeiras sejam susceptíveis a estes fatores relacionados a altitude, os parceiros na simbiose são secundários e não necessariamente seguem o mesmo padrão estabelecido para seus hospedeiros. A colonização micorrízica arbuscular das três espécies não influenciou nas concentrações de macro- e micronutrientes presentes nos tecidos foliares de E. glabriflora, Z. racemosa e A. micrantha, permitindo concluir que os benefícios da associação planta hospedeira e FMA não são necessariamente refletidos na relação PCM e nutrientes nos tecidos foliares das plantas. A ausência de maiores influências das variáveis edáficas, inclusive de nutrientes importantes como P e N, sobre a colonização micorrízica indica que embora as plantas hospedeiras respondam a variáveis ambientais como topografia e textura do solo, os seus parceiros na simbiose não necessariamente seguem os mesmos padrões. As densidades de esporos nas rizosferas de E. glabriflora, Z. racemosa e A. micrantha apresentaram amplas variações ao longo do gradiente topográfico, porém não apresentaram qualquer padrão de distribuição que as relacionasse com a altitude. Os resultados neste estudo indicam que uma variação topográfica de apenas 70 metros na altitude não
66 induziu a variações ambientais suficientemente drásticos para que haja uma esporulação marcante e diferenciada entre os indivíduos da família Leguminosae estudados. Em nenhuma das três espécies foi encontrada qualquer relação entre a densidade de esporos nas rizosferas e as porcentagens de colonização micorrízica presentes nas raízes. Diferentes variáveis das propriedades físicas e químicas influenciaram a densidade de esporos presentes na rizosfera de E. glabriflora, Z. racemosa e A. micrantha sugerindo que possivelmente essas espécies estejam sendo colonizadas por diferentes FMA.
67 REFERÊNCIAS BIBLIOGRÁFICAS Abbot, L.K. & Gazey, C An ecological view of the formation of VA mycorrhizas. Plant and soil, 159: Alencar, A; Nepstad, N; Mcgrath, D; Moutinho, P; Pacheco, P; Diaz, M. D. C. V E Filho, B. S Desmatamento na Amazônia: indo além da emergência crônica. Manaus, Instituto de Pesquisa Ambiental da Amazônia (Ipam), 89 p. Allen, M.F The Ecology of Mycorrhizae. Cambrigde University Press, Cambridge. Allen, O.N. & Allen, E.K The leguminosae: a source book of characteristics, uses and nodulation. Madison, The University of Wisconsin Press, 812p. Anderson, R.C.; Hetrick, B.A.D.; Wilson, G.W.T Mycorrhizal dependence of Andropogon gerardii and Schizachyrium scoparium in two prairie soils. Am. Midl. Nat., 132: Austin, M.P The potential contribution of vegetation ecology to biodiversity research. Ecography, 22: Barcelar-Lima, C.G.; Freire, D.C.B.; Coletto-Silva, A.; Costa, K.B.; Laray, J.P.B.; Vilas-Boas, H.C.; Carvalho-Zilse, G.A Melitocoria de Zygia racemosa (Ducke) Barneby & Grimes por Melipona seminigra merrillae Cockerell, 1919 y Melipona compressipes manaosis Schwars, 1932 (Hymenoptera, Meliponina) en la Amazônia Central, Brasil. Acta Amaz., 36(3): Becker, B.K Geopolítica da Amazônia. Est. Avan., 19(53): Bellei, M.M. & Carvalho, E.M Ectomicorrizas. In: Cardoso, E.J.B.N.; Tsai, S.M.; Neves, M.C.P (cords.) Microbiologia do Solo. Campinas: Sociedade Brasileira de Ciência do Solo. p Beltrati, C.M Morfologia e anatomia de sementes. Pós-Graduação em Ciências Biológicas. Área de Biologia Vegetal. Departamento de Botânica. Instituto de Biociências, UNESP. Rio Claro, SP. 112p. Benjamin, P.K.; Anderson, R.C.; Liberta, A.E Vesicular-arbuscular mycorrhizal ecology of little bluestem across a prairie-forest gradient. Can. J. Bot., 67:
68 Bethlenfalvay, G.J.; Ulrich, J.M.; Brown, M.S Planta response to mycorrhizal fungi: host, endophyte and soil effects. Soil Sci. Soc. Am. J., 49: Bever, J.D.; Morton, J.; Antonovics, J.; Schultz, P.A Host-dependent sporulation and species diversity of arbuscular mycorrhizal fungi in a mown grassland. J. of Ecol., 84(1): Bever, J.D Host-especifity of AM fungal population grouwth rates can generate feedback on plant growth. Plant and Soil, 244(1/2): Bonetti, R.; Oliveira, L.A.; Magalhães, F.M.M População de rhizobium spp. e ocorrência de micorriza VA em cultivos de essências florestais. Pesq. Agropec. Bras., 19 (s/n): Braga-Neto, R Diversidade e padrões de distribuição espacial de fungos de liteira sobre o solo em floresta de terra firme na Amazônia Central. Dissertação de mestrado, INPA/UFAM, Manaus, Amazonas. 271 p. Brundrett, M.C Mycorrhizas in natural ecossystems. Adv. Ecol. Res., 27: Brundrett, M. C., Bougher, N., Grove, T. E Malajczuk, N Working with Mycorrhizas in Forestry and Agriculture. ACIAR (Australian Centre for International Agriculture Research) Monograph 32, Australia. Caris, C.; Hördt, W.; Hawkins, H.J.; Römheld, V.; George, E Studies of iron by arbuscular mycorrhizal hyphae grom soil to peanut and sorghum plants. Mycorrhiza, 8: Carneiro, M.S.C.; Siqueira, J.O.; Moreira, F.M.S.; Carvalho, D.; Botelho, S.A.; Junior, O. J.S Micorriza arbuscular em espécies arbóreas e arbustivas nativas de ocorrência no sudeste do Brasil. Cerne, 4(1): Carrenho, R.; Trufem, S.F.B.; Bononi, V.L.R Fungos micorrizícos arbusculares em rizosfera de três espécies de fitobiontes instaladas em áreas de mata ciliar revegetada. Acta Bot. Bras., 15(1): Carvalho, L.M.; Correia, P.M.; Ryel, R.J.; Martins-Loução, M.A Spatial variability od arbuscular mycorrhizal fungal spores in two natural plant communities. Plant and Soil, 251: Castilho, C.V.; Magnusson, W.E.; Araújo, R.N.O.; Luizão, R.C.C.; Luizão, F.J.; Lima, A.P.; Higuchi, N variation in aboveground tree live biomass
69 in a central Amazonian Forest: Effects of soil and topography. For. Ecol. and Man., 234: Chauvel, A.; Lucas, Y.; Boulet, R On the genesis of the soil mantle of the region of Manaus, Cantral Amazonia, Brazil. Experientia, 43: Claridge, A.W., Barry, S.C., Cork, S.J., Trappe, J.M Diversity and habitat relationships of hypogeous fungi. II. Factors influencing the occurrence and munber of taxa. Biod. and Cons., 9: Clark, D.B. & Clark, D.A Landscape-scale variation in forest structure and biomass in a tropical rain forest. For. Ecol. and Man., 137: Clark, R.B. & Zeto, S.K Mineral acquisition by mycorrhizal maize grown on acid and alkaline soil. Soil Biol. Biochem., 28: Classen, V.P.; Zasoski, R.J A containerised staining system for mycorrhizal roots. New Phytol., 121: Colozzi-Filho, A. & Siqueira, J.O Micorrizas vesículo-arbusculares em mudas de cafeeiro. I. Efeito da inoculação e adubação fosfatada no crescimento e nutrição. R. Bras. Ci. Solo, 10: Corby, H.D.L The systematic value of leguminous root nodules. In: Polhill, R.M. & Haven, P.H. (eds.). Advances in Legume Systematics. Part. 2. Kew, Royal Botanic Gardens, p Costa, F. R. C; Magnusson, W. E; Luizão, R. C. C Mesoscale distribution patterns of Amazonian understorey herbs in relation to topography, soil and watersheds. J. of Ecol., 93: Costa, F.R.C Mesoscale gradients of herb richness and abundance in Central Amazonia. Biotropica, 38(6): Couto, L.B Aspectos sazonais da colonização micorrízica e fatores de fertilidade do solo em plantas de acerola Malphigia sp, café Coffea arabica e laranja Citrus sp em um latossolo do município de Manaus/AM. Dissertação de Mestrado. Manaus: INPA/UFAM.62p. Cuenca, G.; Andrade, Z.; Lovera, M.; Fajardo, L.; Meneses, E Mycorrhizal response of Clusia pusilla growing in two different soils in the field. Trees, 17: Daniel, B.A. & Skipper, H.D Methods of the recovery and quantitative estimation of propagules from soil. In: Methods and principles of Mycorrhizal Research. N.C. Schenck, American Phitopathological Society, St. Paul. P
70 Daws, M.I.; Pearson, T.H.H.; Burslem, D.F.R.P.; Mullins, C.E.; Dalling, J.W Effects of topographic position, leaf litter and seed size on seedling demography in a semi-deciduous tropical forest in Panamá. Plant Ecol., 179: Di Stasi, L.C. & Himura-Lima, C.A Plantas medicinais na Amazônia e na Mata Atlântica. UNESP. 2ed. São Paulo, SP. 604p. Eira, A.F.; Carvalho, P.C.T A decomposição da matéria orgânica pelos microrganismos do solo e sua influência nas variações do ph. Rev. de Agri., 45: EMBRAPA Manual de Análises Químicas de Solos, Plantas e Fertilizantes. Embrapa Solos, Embrapa Informática Agropecuária, Brasília. 370p. Englander, L Endomycorrhizae by septate fungi. In: Schenck, N.C. (eds) Methods and principles of mycorrhizal research. Am. Phytopathol. p Escudero, V. & Mendonza, R Seasonal variation of arbuscular mycorrhizal fungi in temperate grasslands along a wide hydrologic gradient. Mycorrhiza, 15: Fisher, M.A. & Fulé, P.Z Changes in forest vegetation and arbuscular mycorrhizae along a steep elevation gradient in Arizona. For. Ecol. and Man., 200: Freitas, R.O Associações entre fungos micorrízicos arbusculares e espécies pioneiras em capoeiras na Amazônia Central. Dissertação de mestrado. Manaus: INPA/UFAM. 80 p. Friese, C.F. & Koske, R.E The spatial dispersion of spores of vesiculararbuscular mycorrhizal fungi in a sand dune: microscale patterns associated with the root architecture of American beachgrass. Mycol. Res., 95: Frioni, L.; Minasian, H.; Volfovicz, R Arbuscular mycorrhizae and ectomycorrhizae in native tree legumes in Uruguay. For. Ecol. and Man., 115: Gerdemann, J.W.; Nicolson, T.H Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans. of the Brit. Mycol. Soc., 46: Grandcourt, A.; Epron, D.; Montpied, P.; Louisanna, E.; Bereau, M.; Garbaye, J.; Guehl, J. M Contrasting responses to mycorrhizal inoculation
71 and phosphorus availability in seedlings of two tropical rainforest tree species. New Phytogist., 161: Gross, E.; Cordeiro, L.; Caetano, F.H Nodulação e micorrização em Anadenanthera peregrina var. falcata em solo de cerrado autoclavado e não autoclavado. R. Bras. Ci. Solo, 28: Grove, T.S.; Le Tacon, F Mycorrhizal in plantation forestry. Adv. Plant Pathol., 9: Guitton, T.L Ocorrência de micorrizas vesículo arbusculares em algumas espécies florestais da Amazônia com as características físicas e químicas do solo e estado nutricional das plantas. Dissertação de Mestrado. Manaus: INPA/UFAM. 144p. Harley, J.L. & Smith, S.E Mycorrhizal symbiosis. London, Academic Press. 438p. Hart, M.M.; Reader, R.J.; Klironomos, J.N Life-history strategies of arbuscular micorrhizal fungi in relation to their sucessional dynamic. Mycologia, 93(6): Honrubia, M.; Torres, P.; Diaz, G.; Cano, A Manual para micorrizar plantas em viveiros Forestales. Instituto Nacional para la Conservación de la Naturaleza. Proyecto Lucdeme VI Monografia 54. ICONA, España, p. 43. Instituto Nacional de Pesquisas Espaciais (INPE). Monitoramento da Floresta Disponível em: < Acesso em: 15 jan INVAM. International culture collection of vesicular and arbuscular mycorrhizal fungi. Species Description. Disponível em:< Acesso em: 26 jun Islam, R. & Ayanaba, A effect of seed inoculation and pre-infecting cowpea (Vignia unguiculata) with Glomus mosseae on growth and seed yield of the plants under field conditions. Plant and Soil, 61: Isobe, K.; Aizawa, E.; Iguchi, Y.; Ishii, R Distribution of arbuscular mycorrhizal fungi in upland field soil of Japan, 1. Relationship between spore density and the soil environmental factor. Plant Prod. Sci., 10(1): Jackson, R.M.; Mason, P.A Mycorrhiza. The Institute of Biology s Studies in Biology no Edward Arnold.
72 Johnson, N.C.; Zak, D.R.; Tilman, D.; Pfleger, P.L Dynamics of vesiculararbuscular mycorrhizae during old field sucession. Oecologia, 86: Johnson, N.C.; Graham, J.H.; Smith, F.A Functioning of mycorrhizal associations along the mutualism parasitism continuum. New Phytol., 135: Johnson, N.C.; Wolf, J.; Reyes, M.A.; Panter, A.; Koch, G.W.; Redman, A Species of plants and associated arbuscular mycorrhizal fungi mediate mycorrhizal response to CO2 enrichment. Glob. Chan. Biol., 11: Jones, M.D.; Durall D.M.; Tinker, P.B A comparison of arbuscular and ectomycorrhizal Eucalyptus coccifera: growth response, phosphorus uptake efficiency and external hyphal production. New Phytol., 140: Kellman, M. & Tackaberry, R Tropical environments: the functioning and management of tropical ecosystems. London: Routledge, 380 p. Kinupp, V.F. & Magnusson, W.E Spatial patterns in the understorey shrub genus Psychotria in central Amazonia: effects of distance and topography. J. of Trop. Ecol., 21: Kitamura, P.C. A Amazônia e o desenvolvimento sustentável Brasília: EMBRAPA SPI, 182p. Krügner, T.L.; Tomazello Filho, M Ocorrência de micorrizas em espécies de Pinus e identificação dos fungos associados. Boletim Técnico. Instituto de Pesquisas e Estudos Florestais, Piracicaba p. 7. Larcher, W Ecofisiologia Vegetal. São Carlos: RIMA, 531p. Laurance, W.F.; Fearnside, P.M.; Laurance, S.G.; Delamonica, P.; Lovejoy, T.E.; Merona, J.M.R.; Chambers, J.Q.; Gascon, C Relationship between soils and Amazon forest biomass: a landscape-scale study. For. Ecol. and Man., 118: Leal, P.L Fungos micorrízicos arbusculares isolados em culturas armadilhas de solos sob diferentes sistemas de uso na Amazônia. Dissertação de mestrado. Lavras: UFLA, 67 p. Lekberg, Y.; Koide, R.T.; Rohr, J.R.; Aldrich-Wolfe, L.; Morton, J.B Role of niche restrictions and dispersal in the composition of arbuscular mycorrhizal fungal communities. J. of Ecol., 95:
73 Lohman, M.L Occurrence of mycorrhiza in Iowa forest plants. University of Iowa Studies in Natural History, 11: Lorenzi, H Árvores Brasileiras. Manual de identificação e cultivo de plantas arbóreas nativas do Brasil. Editora Plantarum. Nova Odessa: SP. Brasil. 327p. Luizão, R.C.C.; Luizão, F.J.; Paiva, R.Q.; Monteiro, T.F.; Sousa, L.S.; Keuijt, B Variation of carbon and nitrogen cycling processes along a topographic gradient in a cantral Amazonian forest. Glob. Chan. Biol., 10: Magnusson, W.E.; Lima, A.P.; Luizão, R.C.C.; Luizão, F.; Costa, F.R.C.; Castilho, C.V.; Kinupp, V.F RAPELD: a modification of the Gentry method for biodiversity surveys in long-term ecological research sites. Biota Neotropica, 5(2): 1-6. Maia, L.C.; Trufem, S.F.B Fungos micorrízicos vesículo-arbusculares em solos cultivados no Estado de Pernambuco, Brasil. Rev. Bras. de Bot., 13(1): Malavolta, E Elementos de nutrição mineral de plantas. Piracicaba: Ceres, 215p. Marenco, R.A. & Lopes, N.F Fisiologia Vegetal. Viçosa: UFV, 451p. Margules, C.R.; Pressey, R.L.; Williams, P.H Representing biodiversity: data and procedures for identifying priority areas for conservation. J. of Biosc., 27: Martin, J.; Bereau, M.; Louisanna, E.; Ocampo, J.A Arbuscular mycorrhizas in Dicorynia guianensis and Eperua falcata trees from primary tropical rain Forest of French Guiana. Symbiosis, 31(4): Mcgonigle, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi. New Phytol., 115: Mendonza, R.; Goldman, V.; Rivas, J.; Escudero, V.; Pagani, E.; Collantes, M.; Marbán, L Poblaciones de hongos micorrízicos arbusculares em relación com propriedades Del suelo y planta hospedante en pastizales de Tierra del Fuego. Ecol. Austr., 12: Mertens, J The characterization of selected physical and chemical soil properties of the surface soil layer in the Reserva Ducke, Manaus,
74 Brazil, with emphasis on their spatial distribution. Bachelor Thesis. Humboldt-Universität Zu Berlin. Berlin. 46 pp. Miranda, J.C.C. & Miranda, L.N Seleção e Recomendação de uso de espécies de fungos micorrizícos arbusculares. Comunicado Técnico Embrapa Cerrados, Planaltina. 52: 1-3. Mohmmad, M.J.; Hamad, S.R.; Malkawi, H.I Population of arbuscular mycorrhizal fungi in semi-arid environment of Jordan as influenced by biotic and abiotic factors. J. of Arid Envir., 53: Moreira, F.W.; Oliveira, L.A.; Becker, P Ausência de micorrizas vesículoarbusculares efetivas em Lecythidaceas numa área de floresta primária da Amazônia Central. Acta Amaz., 27(1):3-8. Moreira, F.M.S. & Siqueira, J.O Microbiologia e bioquímica do solo. Lavras: Editora UFLA. 626p. Morton, J.B Taxonomy of VA mycorrhizal fungi: classification, nomenclature, and identification. Mycotaxon, 32: Morton, J.B.; Bentivenga, S.P.; Wheeler, W.W Germplasm in the international collection of arbuscular and vesicular-arbuscular mycorrhizal fungi (INVAM) and producers for culture development, documentation and storage. Mycotaxon, 48: Morton, J.B. & Redcker. Archaeosporaceae and Two new Paraglomaceae, families with tow of Glomales, new genera Archaeospora and Paraglomus, based on concordant molecular and morphological characters. Mycologia, 93: Oliveira, L.A.; Moreira, F.W., Moreira, F.M.S Ocorrência de microrganismos benéficos em ecossistemas Amazônicos. In: Noda, H.; Souza, L.A.G.; Fonseca, O.J.M. (Eds.). Duas décadas de contribuições do INPA à pesquisa agronômica no trópico úmido. Instituto de Pesquisas da Amazônia, Manaus. pp Oliveira, L.A.; Guitton, L.T.; Moreira, F.W Relações entre as colonizações por fungos micorrizícos arbusculares e teores de nutrientes foliares em oito espécies florestais da Amazônia. Acta Amaz., 29: Oliveira, A.N.; Oliveira, L.A; Figueiredo, A.N Colonização micorrízica e concentração de nutrientes em três cultivares de bananeiras em um latossolo amarelo da Amazônia Central. Acta Amaz., 33(3):
75 Pande, M.; Tarafdar, J.C Arbuscular mycorrhizal fungal diversity in neem-based agroforestry systems in Rajasthan. Appl. Soil Ecol., 26: Patreze, C.M. & Cordeiro, L Nitrogen-fixing and vesicular-arbuscular mycorrhizal symbioses in some tropical legume trees of tribe Mimoseae. For. Ecol. and Man., 196: Peng, S.; Eissenstat, D.M.; Graham, J.H.; Williams, K.; Hodge, N.C Growth depression in mycorrhizal citrus at high-phosphorus supply. Analysis of carbon costs. Plant Physiol., 101: Pires, J.M. The Forest Ecosystems Of Brazilian Amazon Apud: Moran, E.F. A Ecologia Humana das Populações da Amazônia. Petrópolis: Vozes, Phillips, J.M. & Hayman, D.S Improved producers for clearing roots and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Transactions of the British Mycological Society, 55 (1): Raij, B. van Fertilidade do solo e adubação. Piracicaba: Agronômica Ceres, Associação Brasileira para a Pesquisa da Potassa e do Fosfato, 343p. Ranzani, G Identificação e caracterização de alguns solos da Estação Experimental de Silvicultura Tropical. Acta Amazônica, 10(1): Raven, P.H.; Evert, R.F.; Eichhom, S.E Biologia Vegetal. Rio de Janeiro: Guanabara Koogan S.A. 906p. Ribeiro, J.E.L.S.; Hopkins, M.J.G.; Vicentini, A.; Sothers, C.A.; Costa, M.A.S.; Brito, J.M.; Souza, M.A.D.; Matins, L.H.P.; Lohmann, L.G.; Assunção, P.A.C.L.; Pareira, E.C.; Silva, C.F.; Mesquita, M.R.; Procópio, L Flora da Reserva Ducke: Guia de Identificação das Plantas Vasculares de uma Floresta de Terra Firme na Amazônia Central. INPA/DFID, Manaus, Brasil. Robson, A.D. & Abbott, L.K The effect of soil acidity on mycrobial activity in soils. In: Robson, A.D. (ed.). Soil acidity and plant growth. Academic Press, Sydney, 99: Ropero, C.A.L Micorrizas arbusculares e assimilação de nutrientes em uma meso-escala na região Amazônica. Dissertação de mestrado. Manaus: INPA/UFAM. 49 p.
76 Sanchez, P.; Bandy, P.E.; Villachica, J.H.; Nicolaides Amazon Basin Soils: Management for Continuous Crop Production. Science, 216: Schachtman, D.P.; Reid, R.J.; Ayling, S.M Phosphorus uptake by plants from soil to cell. Plant Physiol., 116: Schultz, P.A.; Miller, R.M.; Jastrow, J.A.; Rivetta, C.V.; Bever, J.D Evidence of a mycorrhizal mechanism for the adaptation of Andropogon gerardii (Poaceae) to high- and low-nutrient prairies. Am. J. Bot., 88: Schussler, A; Schwarzott, D.; Walker, C A new fungal phylum the Glomeromycota: phylogenyand evolution. Mycology Research, 105(12): Sieverding, E Vesicular-arbuscular mycorrhiza management in tropical agrosystems. Alemanha: Eschborn, 371 p. Silva, E.M.R.; Franco, A.A.; Dias, L.E.; Campello, E.F.C Fungos micorrízicos em leguminosas arbóreas revegetando solo degradado. In: Universidade Federal de Santa Catarina. Reunião Brasileira sobre Micorrizas (5ª, Florianópolis, SC), Resumos, pp. 25. Silveira, A.P.D Micorrizas. In: Cardoso, E.J.B.N.; Tsai, S.M.; Neves, M.C.P (cords.) Microbiol. do Solo. Campinas: Sociedade Brasileira de Ciência do Solo. p Siqueira, J.O Micorrizas: forma e função. In: Reunião Brasileira sobre Micorrizas, Lavras. Anais...Lavras: FAEPE, Siqueira, J.O.; Colozzi-Filho, A.; Oliveira, E.; Ocorrência de micorrizas vesicular-arbuscular em agro e ecossistemas do estado de Minas Gerais. Pesq. Agrop. Bras., 24(12): Siqueira, J.O Micorrizas arbusculares. In: Araújo, R.S. & Hungria, M. (eds). Microrganismos de importância agrícola. Brasília, DF, EMBRAPA SPI, p Siqueira, J.O.; Carneiro, M.A.C.; Curi, N.; Rosado, S.C.S; Davide, A.C Mycorrhizal colonization and mycotrophic growth of native woody species as related to sucessional groups in Southeastern Brazil. For. Ecol. Manag., 107:
77 Siqueira, J.O.; Saggin-Júnior, O.J Dependency on arbuscular mycorrhizal fungi and responsiveness of some Brazilian native woody species. Mycorrhiza, 11: Siqueira, J.O.; Lambais, M.R.; Stürmer, S.L Fungos micorrízicos arbusculares. Biotec. Ci. & Desen., 25: Smith, E.S.; Read, J.D Mycorrhizal Symbiosis. 2 ed. New York: Academic Press. 605p. Sombroek, W.G References Bibliografia sobre os solos da Amazônia. In:. Amazon Soils. Wageningen, C.A.P.D. Souza, L.A.G. Cientistas estudam produtos renováveis das leguminosas arbóreas da Amazônia. Disponível em: < 183>. Acesso em: 24 mar St. John, T Uma lista de espécies de plantas tropicais brasileiras naturalmente infectadas com micorriza vesicular-arbuscular. Acta Amaz., 10(1): St. John, T.V. & Uhl, C Mycorrhizae in the rain forest at San Carlos de Rio Negro, Venezuela. Acta Cient. Venezoelana, 34: Taiz, L.; Zeiger, E Fisiologia Vegetal. Porto Alegre: ARTMED, 719p. Takyu, M.; Aiba, S.I.; Kitayama, K Changes in biomass, productivity and decomposition along topographical gradients under different geological condiotions in tropical lower montane Forest on Mount Kinabalu, Borneo. Oecologia, 134: Tinker, P.B.H Effects of vesicular-arbuscular mycorrhiza on higher plantas. Symp. Soc. Expo. Biol., 29: Trappe, J.M Phylogenetic and ecological aspects os mycotrophic in the Angiosperms from an evolutionary satandpoint. In: Safir, G.R.; (Ed.). Ecophysiology of VA Mycorrhizal Plants. Boca Raton. p Tuomisto, H.; Ruokolainen, K.; Yli-Halla, M Dispersal, environment and floristic variation of western Amazonia forests. Science, 299: Vasconcelos, I.; Almeida, R.T.; Mendes, P.F Ocorrência de rizóbios e endomicorrizas em leguminosas arbóreas e arbustivas do Estado do Ceará, Brasil. Cs. Agron. Fortaleza, 15 (1/2) Vierheilig, H. & Ocampo, J.A Receptivity of various wheat cultivars to infection by VA-mycorrhizal fungus influenced by inoculum potential and
78 the relation of VAM-effectiveness to succinic dehydrogenase activity of the mycelium in the root. Plant Soil, 133: Warcup, J.H. & Talbot, P.H.B Perfect states of Rhizoctonias associated with orchids. New Phytol., 84: Wilcox, H.E Mycorrhizal Associations. In: Nakas, J.P.; Hagedorn, C. (Ed.). Biotechnology of plant-microbe interactions. New York: McGrawHill, Wilkinson, L Systat: The System for Statistic. Systat Inc., Evanston, Illinois. 822 p. Wood, T.E.; Lawrence, D.; Clark, D Determinants of leaf litter nutrient cycling in a tropical rain forest: Soil fertility versus topography. Ecosys., 9: Zangaro, W.; Nisizaki, S.M.A.; Domingos, J.C.B.; Nakano, E.M Mycorrhizal response and successional status in 80 woody species from south Brazil. J. of Trop. Ecol., 19: Zhao, Z.W.; Xia, Y.M.; Qin, X.Z.; Li, X.W.; Cheng, L.Z.; Sha, T.; Wang, G.H Arbuscular mycorrhizal status of plants and the spore density of arbuscular mycorrhizal fungi in the tropical rain Forest of Xishuangbanna, southwest China. Mycorrhiza, 11: APÊNDICE Apêndice 1. Características químicas do solo das parcelas nas rizosferas das
79 três espécies de Leguminosae na RFAD. Com as médias e seus desvios padrão (parênteses)...66 Apêndice 2. Características físicas do solo das parcelas nas rizosferas das três espécies de Leguminosae na RFAD. Com as médias e seus desvios padrão (parênteses)...67 Apêndice 3. Imagens da coleta de raízes (A), das raízes ampliadas e micorrizadas (B) e de esporos de FMA (C)...68 Apêndice 1. Características químicas do solo das parcelas nas rizosferas das três espécies de Leguminosae na RFAD. Com as médias e seus desvios padrão (parênteses).
80 Andira micrantha Zygia racemosa Eperua glabriflora Espécie Parcela LO3T1500 n=5 LO3T2500 n=4 LO3T3500 n=4 LO4T500 n=5 LO4T1500 n=3 LO4T4500 n=5 LO5T500 n=5 LO6T3500 n=5 LO6T4500 n=3 LO7T500 n=4 LO7T3500 n=4 LO7T4500 n=5 LO8T500 n=4 LO8T1500 n=4 LO8T2500 n=4 LO8T3500 n=5 LO3T500 n=4 LO3T4500 n=3 LO4T1500 n=3 LO4T2500 n=5 LO4T3500 n=4 LO5T1500 n=5 LO5T2500 n=4 LO5T3500 n=4 LO5T4500 n=4 LO6T3500 n=3 LO7T1500 n=5 LO8T4500 n=5 LO3T2500 n=4 LO6T500 n=3 LO6T1500 n=4 LO6T2500 n=4 LO6T3500 n=5 LO7T2500 n=3 Altitude (m) 61,7 53,3 55,3 46,7 55,4 98,9 56,0 100,9 75,7 58,3 92,2 100,0 52,5 49,4 63,8 56,2 81,7 88,7 55,4 100,1 105,1 93,6 73,9 103,4 94,8 100,9 84,7 102,2 53,3 64,5 53,5 60,5 100,9 101,5 (H2O) 4,8 N Total -1 g. Kg 1,97 Química do solo Macronutrientes Micronutrientes P Total K Ca Mg Al Fe Zn -1-1 mg. Kg-1 Cmolc. Kg mg. Kg 1,78 25, ,4 6, ,62 (±0,13) (±0,88) (±0,07) ph (±4,2) (±307) (±32,9) (±1,74) (±87) (±3,56) 2+ Mn 6,96 (±4,32) 4,6 1,39 2,33 14, ,3 4, ,30 5,00 (±0,13) (±0,90) (±0,51) (±2,2) (±76) (±14,4) (±1,09) (±109) (±0,45) (±7,46) 4,8 4,38 7,77 24, ,1 4, ,85 6,10 (±0,6) (±0,53) (±3,85) (±5,6) (±89) (±16,0) (±1,23) (±10) (±0,31) (±4,67) 4,5 2,75 3,35 22, ,7 3, ,00 5,58 (±0,20) (±2,29) (±0,28) (±2,7) (±187) (±24,8) (±1,84) (±60) (±4,02) (±3,20) 4,8 1,04 2,50 13, ,0 4, ,56 4,96 (±0,33) (±0,37) (±0,55) (±2,4) (±80) (±14,6) (±0,28) (±99) (±1,62) (±3,70) 4,7 1,88 1,74 24, ,0 6, ,56 4,08 (±0,16) (±1,00) (±0,22) (±10,2) (±192) (±33,7) (±1,31) (±133) (±1,40) (±2,78) 4,6 3,96 10,34 23, ,5 3, ,50 4,20 (±0,38) (±1,24) (±3,39) (±11,3) (±106) (±17,9) (±1,04) (±3) (±0,43) (±1,89) 4,7 2,01 1,54 20, ,9 3, ,64 2,44 (±0,52) (±1,19) (±0,26) (±5,9) (±51) (±5,0) (±1,87) (±95) (±1,4) (±1,02) 4,8 3,94 1,85 14,2 62 9,1 5, ,20 3,76 (±0,20) (±0,66) (±0,06) (±11,8) (±54) (±5,9) (±2,95) (±167) (±1,96) (±2,28) 4,6 3,39 2,05 17, ,0 4, ,37 6,15 (±0,09) (±4,39) (±0,20) (±4,0) (±421) (±46,3) (±0,73) (±51) (±4,51) (±4,03) 5,0 2,34 1,76 29, ,6 3, ,92 7,85 (±0,18) (±0,97) (±0,46) (±5,9) (±103) (±20,1) (±2,31) (±148) (±4,99) (±6,35) 4,7 4,09 1,73 22, ,7 3, ,16 10,44 (±0,15) (±3,26) (±0,43) (±6,6) (±149) (±21,5) (±1,97) (±129) (±3,76) (±12,93) 4,7 8,33 3,62 24, ,7 5, ,82 6,9 (±0,22) (±8,48) (±0,56) (±3,1) (±78) (±12,6) (±2,59) (±94) (±3,07) (±4,57) 4,8 4,51 5,16 26, ,6 3, ,27 7,02 (±0,45) (±4,39) (±1,52) (±7,3) (±293) (±18,6) (±1,09) (±155) (±4,11) (±2,89) 4,7 4,34 3,73 21, ,7 2, ,95 5,97 (±0,06) (±1,47) (±2,19) (±9,5) (±183) (±22,5) (±2,00) (±144) (±4,53) (±4,01) 4,9 5,23 5,03 17, ,9 4, ,14 5,72 (±0,13) (±3,21) (±0,75) (±3,1) (±298) (±45,5) (±2,56) (±103) (±1,29) (±2,82) 4,6 3,87 2,04 18, ,0 3, ,55 10,52 (±0,15) (±4,17) (±0,10) (±6,7) (±81) (±9,1) (±1,53) (±15) (±2,24) (±13,11) 4,7 2,38 1,76 27, ,3 6, ,03 4,46 (±0,18) (±0,52) (±0,32) (±7,2) (±70) (±6,3) (±3,08) (±170) (±0,92) (±4,12) 4,7 1,10 7,81 21, ,6 3, ,63 8,43 (±0,07) (±0,41) (±3,60) (±6,8) (±209) (±21,9) (±2,30) (±102) (±2,49) (±4,28) 4,7 2,67 2,11 39, ,3 1, ,88 4,00 (±0,44) (±1,06) (±0,33) (±11,0) (±86) (±33,1) (±0,67) (±36) (±0,31) (±2,80) 4,9 2,65 1,59 26, ,8 5, ,25 3,92 (±0,15) (±0,93) (±0,02) (±11,3) (±21) (±13,7) (±2,89) (±207) (±2,28) (±3,21) 4,8 1,52 2,51 18, ,0 5, ,00 2,14 (±0,57) (±0,72) (±0,40) (±15,6) (±78) (±12,8) (±1,82) (±62) (±1,53) (±1,27) 4,6 3,19 2,69 20, ,5 3, ,75 5,55 (±0,21) (±3,69) (±0,65) (±4,6) (±226) (±23,6) (±2,69) (±129) (±4,48) (±3,77) 4,8 2,55 1,83 27, ,5 4, ,92 3,22 (±0,18) (±0,34) (±0,23) (±5,6) (±83) (±28,0) (±2,85) (±141) (±0,32) (±2,53) 4,2 2,14 1,58 17, ,7 4, ,40 2,70 (±0,27) (±0,75) (±0,32) (±9,2) (±178) (±27,9) (±3,06) (±94) (±1,79) (±1,50) 4,8 3,27 1,47 19, ,8 5, ,50 2,10 (±0,14) (±0,54) (±0,19) (±9,6) (±471) (±27,2) (±2,07) (±223) (±1,40) (±1,21) 4,7 2,21 1,33 24, ,0 4, ,00 10,48 (±0,33) (±1,48) (±0,07) (±10,6) (±262) (±41,6) (±2,41) (±109) (±1,51) (±6,48) 4,7 2,91 1,58 24, ,1 4, ,40 4,28 (±0,15) (±1,28) (±0,21) (±8,5) (±193) (±23,4) (±2,99) (±141) (±1,23) (±2,85) 4,7 4,26 2,22 33, ,6 3, ,10 4,35 (±0,07) (±0,19) (±0,16) (±2,8) (±223) (±88,8) (±1,07) (±47) (±0,42) (±3,37) 4,7 2,80 2,96 14, ,6 3, ,03 3,10 (±0,21) (±0,60) (±0,26) (±3,6) (±150) (±21,9) (±1,41) (±124) (±1,34) (±1,04) 4,8 2,27 2,87 12, ,2 5, ,50 3,62 (±0,23) (±1,19) (±0,81) (±5,1) (±61) (±14,8) (±0,58) (±67) (±2,70) (±3,57) 4,2 3,10 6,00 36, ,2 3, ,72 2,82 (±0,45) (±0,94) (±1,27) (±11,9) (±9) (±11,2) (±1,64) (±4) (±0,49) (±1,25) 4,9 3,18 1,46 24, ,8 4, ,18 5,14 (±0,15) (±0,97) (±0,08) (±11,6) (±176) (±59,7) (±1,75) (±97) (±3,63) (±3,03) 4,8 2,97 2,12 26, ,8 5, ,20 3,40 (±0,20) (±2,16) (±0,44) (±4,4) (±134) (±58,6) (±0,90) (±112) (±2,64) (±1,67)
81 Apêndice 2. Características físicas do solo das parcelas nas rizosferas das três espécies de Andira micrantha Zygia racemosa Eperua glabriflora Espécie Leguminosae na RFAD. Com as médias e seus desvios padrão (parênteses). Parcela LO3T1500 n=5 LO3T2500 n=4 LO3T3500 n=4 LO4T500 n=5 LO4T1500 n=3 LO4T4500 n=5 LO5T500 n=5 LO6T3500 n=5 LO6T4500 n=3 LO7T500 n=4 LO7T3500 n=4 LO7T4500 n=5 LO8T500 n=4 LO8T1500 n=4 LO8T2500 n=4 LO8T3500 n=5 LO3T500 n=4 LO3T4500 n=3 LO4T1500 n=3 LO4T2500 n=5 LO4T3500 n=4 LO5T1500 n=5 LO5T2500 n=4 LO5T3500 n=4 LO5T4500 n=4 LO6T3500 n=3 LO7T1500 n=5 LO8T4500 n=5 LO3T2500 n=4 LO6T500 n=3 LO6T1500 n=4 LO6T2500 n=4 LO6T3500 n=5 LO7T2500 n=3 Altitude (m) 61,7 53,3 55,3 46,7 55,4 98,9 56,0 100,9 75,7 58,3 92,2 100,0 52,5 49,4 63,8 56,2 81,7 88,7 55,4 100,1 105,1 93,6 73,9 103,4 94,8 100,9 84,7 102,2 53,3 64,5 53,5 60,5 100,9 101,5 Unidade 9.45 Física do solo Granulometria Areia Silte Argila (%) (±1,82) (±1,34) (±5,96) 0.60 (±2,41) (±10,21) (±2,20) (±1,34) (±1,86) (±3,91) (±2,05) (±9,53) (±24,89) (±2,04) (±1,68) (±1,19) (±6,04) (±3,14) (±1,53) (±2,30) (±9,53) (±1,19) (±8,23) (±28,48) (±2,16) (±12,13) (±1,34) (±4,88) (±5,58) (±3,37) (±52,32) (±2,11) (±2,13) (±3,08) (±2,38) (±9,53) (±4,85) (±1,95) (±19,03) (±2,16) (±1,68) (±1,65) (±1,38) (±2,11) (±2,04) (±4,29) (±17,73) (±3,34) (±1,68) (±1,19) (±1,21)


,")





82 Apêndice 3. Imagens da coleta de raízes (A), raízes ampliadas e micorrizadas (B) e de esporos de FMA (C). A2 A1 A3 A4 hf hf B1 B2 ar ar ve B3 hf B4



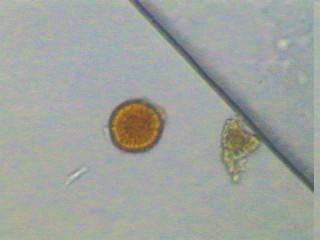
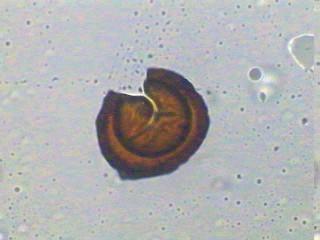
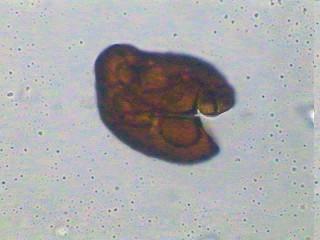
83 C1 C2 C3 C4 C5 C6 Legenda: hf - hifas, ve - vesículas, ar arbúsculos.
Orientador Newton Stamford - UFRPE
 UNIVERSIDADE FEDERAL RURAL DE PERNAMBUCO DEPARTAMENTO DE AGRONOMIA PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIA DO SOLO SEMINÁRIOS EM CIÊNCIA DO SOLO ECOLOGIA DE FUNGOS MICORRíZICOS ARBUSCULARES Rosemberg de V.
UNIVERSIDADE FEDERAL RURAL DE PERNAMBUCO DEPARTAMENTO DE AGRONOMIA PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIA DO SOLO SEMINÁRIOS EM CIÊNCIA DO SOLO ECOLOGIA DE FUNGOS MICORRíZICOS ARBUSCULARES Rosemberg de V.
AS RELAÇÕES ENTRE MACRONUTRIENTES E MICRONUTRIENTES E A FERTILIDADE DO SOLO Pedro Lopes Ferlini Salles Orientadora: Marisa Falco Fonseca Garcia
 AS RELAÇÕES ENTRE MACRONUTRIENTES E MICRONUTRIENTES E A FERTILIDADE DO SOLO Pedro Lopes Ferlini Salles Orientadora: Marisa Falco Fonseca Garcia Coorientador: Flávio Ferlini Salles RELEVÂNCIA O solo é importante
AS RELAÇÕES ENTRE MACRONUTRIENTES E MICRONUTRIENTES E A FERTILIDADE DO SOLO Pedro Lopes Ferlini Salles Orientadora: Marisa Falco Fonseca Garcia Coorientador: Flávio Ferlini Salles RELEVÂNCIA O solo é importante
O SOLO COMO F0RNECEDOR DE NUTRIENTES
 O SOLO COMO F0RNECEDOR DE NUTRIENTES LIQUIDA (SOLUÇÃO DO SOLO) ÍONS INORGÂNICOS E ORGÂNICOS/MICROPOROS SÓLIDA - RESERVATORIO DE NUTRIENTES - SUPERFÍCIE QUE REGULA A CONCENTRAÇÃO DOS ELEMENTOS NA SOLUÇÃO
O SOLO COMO F0RNECEDOR DE NUTRIENTES LIQUIDA (SOLUÇÃO DO SOLO) ÍONS INORGÂNICOS E ORGÂNICOS/MICROPOROS SÓLIDA - RESERVATORIO DE NUTRIENTES - SUPERFÍCIE QUE REGULA A CONCENTRAÇÃO DOS ELEMENTOS NA SOLUÇÃO
MATÉRIA ORGÂNICA DO SOLO (MOS) Fertilidade do Solo Prof. Josinaldo
 Fertilidade do Solo Prof. Josinaldo") MATÉRIA ORGÂNICA DO SOLO (MOS) Fertilidade do Solo Prof. Josinaldo 1 ASPECTOS GERAIS - MOS todos os compostos orgânicos do solo - Influência os atributos do solo - Teor no solo amplamente variável (0,5
MATÉRIA ORGÂNICA DO SOLO (MOS) Fertilidade do Solo Prof. Josinaldo 1 ASPECTOS GERAIS - MOS todos os compostos orgânicos do solo - Influência os atributos do solo - Teor no solo amplamente variável (0,5
APRESENTAÇÃO: FERTILIZANTE TERRAPLANT
 APRESENTAÇÃO: FERTILIZANTE TERRAPLANT DESCRIÇÃO DO PRODUTO Fertilizante proveniente de cama de aviário; Fonte de macro e micro nutrientes; Fonte excepcional de matéria orgânica. DESCRIÇÃO DO PRODUTO Para
APRESENTAÇÃO: FERTILIZANTE TERRAPLANT DESCRIÇÃO DO PRODUTO Fertilizante proveniente de cama de aviário; Fonte de macro e micro nutrientes; Fonte excepcional de matéria orgânica. DESCRIÇÃO DO PRODUTO Para
FACULDADE VÉRTICE CURSO AGRONOMIA MICROBIOLOGIA DO SOLO TEMAS: BIOTA E AGREGAÇÃO DO SOLO E OS PRINCIPAIS MICROORGANISMOS DE IMPORTÂNCIA AGRÍCOLA
 FACULDADE VÉRTICE CURSO AGRONOMIA MICROBIOLOGIA DO SOLO TEMAS: BIOTA E AGREGAÇÃO DO SOLO E OS PRINCIPAIS MICROORGANISMOS DE IMPORTÂNCIA AGRÍCOLA PROFESSORA: MARIA LITA P. CORREA EVOLUÇÃO DO SOLOS Cianob,
FACULDADE VÉRTICE CURSO AGRONOMIA MICROBIOLOGIA DO SOLO TEMAS: BIOTA E AGREGAÇÃO DO SOLO E OS PRINCIPAIS MICROORGANISMOS DE IMPORTÂNCIA AGRÍCOLA PROFESSORA: MARIA LITA P. CORREA EVOLUÇÃO DO SOLOS Cianob,
Interpretando as relações evolutivas
 Tempo COLÉGIO TIRADENTES DA PMMG Unidade Gameleira ATIVIDADE EM SALA DISCIPLINA: Ciências DATA: ENSINO: Fundamental II ESPECIALISTA: Iris e Carolina PROFESSOR (A): Andrea Lima Alves Ruislan ALUNO (A):
Tempo COLÉGIO TIRADENTES DA PMMG Unidade Gameleira ATIVIDADE EM SALA DISCIPLINA: Ciências DATA: ENSINO: Fundamental II ESPECIALISTA: Iris e Carolina PROFESSOR (A): Andrea Lima Alves Ruislan ALUNO (A):
BIOSFERA E SEUS ECOSSISTEMAS Cap.2
 BIOSFERA E SEUS ECOSSISTEMAS Cap.2 Conceitos Básicos ECOLOGIA Oikos =casa; logos= ciência É a ciência que estuda as relações entre os seres vivos entre si e com o ambiente onde eles vivem Estuda as formas
BIOSFERA E SEUS ECOSSISTEMAS Cap.2 Conceitos Básicos ECOLOGIA Oikos =casa; logos= ciência É a ciência que estuda as relações entre os seres vivos entre si e com o ambiente onde eles vivem Estuda as formas
Biodiversidade e prosperidade económica
 Biodiversidade e prosperidade económica Helena Castro e Helena Freitas Centro de Ecologia Funcional Universidade de Coimbra O que é a biodiversidade? Biodiversidade é a variedade de seres vivos. Aqui se
Biodiversidade e prosperidade económica Helena Castro e Helena Freitas Centro de Ecologia Funcional Universidade de Coimbra O que é a biodiversidade? Biodiversidade é a variedade de seres vivos. Aqui se
Assimilação de Nutrientes Minerais
 Assimilação de Nutrientes Minerais plantas são organismos fotoautoróficos: todos os compostos orgânicos necessários são formados em seu próprio corpo (macro e micro moléculas) a energia necessária para
Assimilação de Nutrientes Minerais plantas são organismos fotoautoróficos: todos os compostos orgânicos necessários são formados em seu próprio corpo (macro e micro moléculas) a energia necessária para
Biologia. Rubens Oda (Julio Junior) Ecologia
 Ecologia") Ecologia Ecologia 1. Observe a cadeia alimentar representada no esquema abaixo. Nomeie o nível trófico no qual é encontrada a maior concentração de energia, indique a letra que o representa no esquema
Ecologia Ecologia 1. Observe a cadeia alimentar representada no esquema abaixo. Nomeie o nível trófico no qual é encontrada a maior concentração de energia, indique a letra que o representa no esquema
Nutrição, Adubação e Calagem
 Nutrição, Adubação e Calagem Importância da nutrição mineral Embora o eucalipto tenha rápido crescimento, este é muito variável. Os principais fatores que interferem no crescimento estão relacionados com
Nutrição, Adubação e Calagem Importância da nutrição mineral Embora o eucalipto tenha rápido crescimento, este é muito variável. Os principais fatores que interferem no crescimento estão relacionados com
Questões de Revisão. 7º ano BIOLOGIA Ana Clara 3º Bimestre
 Questões de Revisão 7º ano BIOLOGIA Ana Clara 3º Bimestre Questão 01) Cladogramas são diagramas que indicam uma história comum entre espécies ou grupos de seres vivos. Os números 3 e 4 no cladograma apresentado
Questões de Revisão 7º ano BIOLOGIA Ana Clara 3º Bimestre Questão 01) Cladogramas são diagramas que indicam uma história comum entre espécies ou grupos de seres vivos. Os números 3 e 4 no cladograma apresentado
Prof. Everlon Cid Rigobelo. Ecologia do Solo
 Prof. Everlon Cid Rigobelo Ecologia do Solo Ecologia do Solo Ubiquidade dos micro-organismos Versatilidade metabólica Tolerância às condições ambientais adversas Fácil dispersão Características intrínsecas
Prof. Everlon Cid Rigobelo Ecologia do Solo Ecologia do Solo Ubiquidade dos micro-organismos Versatilidade metabólica Tolerância às condições ambientais adversas Fácil dispersão Características intrínsecas
Avaliação da velocidade de reação do corretivo líquido na camada superficial de um Latossolo Vermelho distroférrico
 Avaliação da velocidade de reação do corretivo líquido na camada superficial de um Latossolo Vermelho distroférrico Bruna de Souza SILVEIRA 1 ; André Luís XAVIER 1 ; Sheila Isabel do Carmo PINTO 2 ; Fernando
Avaliação da velocidade de reação do corretivo líquido na camada superficial de um Latossolo Vermelho distroférrico Bruna de Souza SILVEIRA 1 ; André Luís XAVIER 1 ; Sheila Isabel do Carmo PINTO 2 ; Fernando
1.1 Conceitos em nutrição de plantas. Outros elementos químicos de interesse na nutrição vegetal.
 1. CONCEITOS 1.1 Conceitos em nutrição de plantas. 1.2 Conceito de nutrientes e critérios de essencialidade. 1.3 Composição relativa das plantas. Outros elementos químicos de interesse na nutrição vegetal.
1. CONCEITOS 1.1 Conceitos em nutrição de plantas. 1.2 Conceito de nutrientes e critérios de essencialidade. 1.3 Composição relativa das plantas. Outros elementos químicos de interesse na nutrição vegetal.
LISTA DE EXERCÍCIOS CIÊNCIAS
 LISTA DE EXERCÍCIOS CIÊNCIAS P1-4º BIMESTRE 6º ANO FUNDAMENTAL II Aluno (a): Turno: Turma: Unidade Data: / /2016 HABILIDADES E COMPETÊNCIAS Compreender o conceito de bioma. Reconhecer fatores bióticos
LISTA DE EXERCÍCIOS CIÊNCIAS P1-4º BIMESTRE 6º ANO FUNDAMENTAL II Aluno (a): Turno: Turma: Unidade Data: / /2016 HABILIDADES E COMPETÊNCIAS Compreender o conceito de bioma. Reconhecer fatores bióticos
Cadeia Produtiva da Silvicultura
 Cadeia Produtiva da Silvicultura Silvicultura É a atividade que se ocupa do estabelecimento, do desenvolvimento e da reprodução de florestas, visando a múltiplas aplicações, tais como: a produção de madeira,
Cadeia Produtiva da Silvicultura Silvicultura É a atividade que se ocupa do estabelecimento, do desenvolvimento e da reprodução de florestas, visando a múltiplas aplicações, tais como: a produção de madeira,
Clima(s) CLIMAS - SOLOS E AGRICULTURA TROPICAL. Mestrado em Direito à Alimentação e Desenvolvimento Rural UC: Agricultura Tropical.
 CLIMAS - SOLOS E AGRICULTURA TROPICAL. Mestrado em Direito à Alimentação e Desenvolvimento Rural UC: Agricultura Tropical.") CLIMAS - SOLOS E AGRICULTURA TROPICAL Mestrado em Direito à Alimentação e Desenvolvimento Rural UC: Agricultura Tropical Óscar Crispim Machado (omachado@esac.pt) ESAC, abril de 2012 Clima(s) Aula 5 Zonas
CLIMAS - SOLOS E AGRICULTURA TROPICAL Mestrado em Direito à Alimentação e Desenvolvimento Rural UC: Agricultura Tropical Óscar Crispim Machado (omachado@esac.pt) ESAC, abril de 2012 Clima(s) Aula 5 Zonas
Edson Vidal Prof. Manejo de Florestas Tropicais ESALQ/USP
 Edson Vidal Prof. Manejo de Florestas Tropicais ESALQ/USP PRINCIPAIS ECOSSISTEMAS CAATINGA 844 Mil Km 2 (9,9%) Território Brasileiro = 9,9% Nordeste = 55,6% Cobertura Atual 30% Fonte: IBAMA 2002 Elaboração:
Edson Vidal Prof. Manejo de Florestas Tropicais ESALQ/USP PRINCIPAIS ECOSSISTEMAS CAATINGA 844 Mil Km 2 (9,9%) Território Brasileiro = 9,9% Nordeste = 55,6% Cobertura Atual 30% Fonte: IBAMA 2002 Elaboração:
Morfologia Vegetal. O corpo da planta
 Morfologia Vegetal O corpo da planta Os sistemas da planta Meristema apical do caule Sistema caulinar Ou axial Crescimento Modular Fitômeros Plasticidade Fenotípica Sistema radicular Ou absortivo-fixador
Morfologia Vegetal O corpo da planta Os sistemas da planta Meristema apical do caule Sistema caulinar Ou axial Crescimento Modular Fitômeros Plasticidade Fenotípica Sistema radicular Ou absortivo-fixador
Fatores de Formação de Solos
 Fatores de Formação de Solos De onde vem o solo? O solo resulta da ação simultânea do clima e organismos que atuam sobre um material de origem (rocha), que ocupa determinada paisagem ou relevo, durante
Fatores de Formação de Solos De onde vem o solo? O solo resulta da ação simultânea do clima e organismos que atuam sobre um material de origem (rocha), que ocupa determinada paisagem ou relevo, durante
Ecossistemas e Saúde Ambiental :: Prof.ª MSC. Dulce Amélia Santos
 11/8/2011 ECOLOGIA Disciplina Ecossistemas E Saúde Ambiental Ecologia é uma área da Biologia que estuda os organismos e suas relações com o meio ambiente em que vivem. Aula 1A: Revendo Conceitos Básicos
11/8/2011 ECOLOGIA Disciplina Ecossistemas E Saúde Ambiental Ecologia é uma área da Biologia que estuda os organismos e suas relações com o meio ambiente em que vivem. Aula 1A: Revendo Conceitos Básicos
Características gerais da Madeira Prof. Dr. Umberto Klock.
 AT073 - Introdução à Engenharia Industrial Madeireira Características gerais da Madeira Prof. Dr. Umberto Klock. 1 Características gerais da Madeira O que iremos apresentar e discutir? Plantas superiores
AT073 - Introdução à Engenharia Industrial Madeireira Características gerais da Madeira Prof. Dr. Umberto Klock. 1 Características gerais da Madeira O que iremos apresentar e discutir? Plantas superiores
Anais do Congresso de Pesquisa, Ensino e Extensão- CONPEEX (2010)
") Anais do Congresso de Pesquisa, Ensino e Extensão- CONPEEX (2010) 5771-5775 AVALIAÇÃO DE GENÓTIPOS DE ARROZ IRRIGADO NO USO DE NITROGÊNIO CARVALHO, Glaucilene Duarte 1 ; DE CAMPOS, Alfredo Borges 2 & FAGERIA,
Anais do Congresso de Pesquisa, Ensino e Extensão- CONPEEX (2010) 5771-5775 AVALIAÇÃO DE GENÓTIPOS DE ARROZ IRRIGADO NO USO DE NITROGÊNIO CARVALHO, Glaucilene Duarte 1 ; DE CAMPOS, Alfredo Borges 2 & FAGERIA,
PLANO DE AULA Nutrição das Plantas Autores: Ana Paula Farias Waltrick, Stephanie Caroline Schubert;
 PLANO DE AULA Nutrição das Plantas Autores: Ana Paula Farias Waltrick, Stephanie Caroline Schubert; 1. DADOS DE IDENTIFICAÇÃO Nível de Ensino: Ensino Médio Ano/Série: 1º ano Disciplina: Biologia 2. TEMA
PLANO DE AULA Nutrição das Plantas Autores: Ana Paula Farias Waltrick, Stephanie Caroline Schubert; 1. DADOS DE IDENTIFICAÇÃO Nível de Ensino: Ensino Médio Ano/Série: 1º ano Disciplina: Biologia 2. TEMA
VII Semana de Ciência Tecnologia IFMG campus
 Avaliação do efeito do corretivo líquido sobre o ph, Ca 2+, Mg 2+, porcentagem de saturação por bases (V) e alumínio (m) em um Latossolo Vermelho distroférrico André Luís Xavier Souza 1 ; Fernando Carvalho
Avaliação do efeito do corretivo líquido sobre o ph, Ca 2+, Mg 2+, porcentagem de saturação por bases (V) e alumínio (m) em um Latossolo Vermelho distroférrico André Luís Xavier Souza 1 ; Fernando Carvalho
Estudo 4 - Oportunidades de Negócios em Segmentos Produtivos Nacionais
 Centro de Gestão e Estudos Estratégicos Ciência, Tecnologia e Inovação Prospecção Tecnológica Mudança do Clima Estudo 4 - Oportunidades de Negócios em Segmentos Produtivos Nacionais Giselda Durigan Instituto
Centro de Gestão e Estudos Estratégicos Ciência, Tecnologia e Inovação Prospecção Tecnológica Mudança do Clima Estudo 4 - Oportunidades de Negócios em Segmentos Produtivos Nacionais Giselda Durigan Instituto
Matéria Orgânica do Solo. Everlon Cid Rigobelo
 Matéria Orgânica do Solo Everlon Cid Rigobelo 1 Solo receptáculo final Recebedor Resíduos orgânicos de origem Vegetal, animal Produtos de suas transformações 2 Solo receptáculo final Vegetação Responsável
Matéria Orgânica do Solo Everlon Cid Rigobelo 1 Solo receptáculo final Recebedor Resíduos orgânicos de origem Vegetal, animal Produtos de suas transformações 2 Solo receptáculo final Vegetação Responsável
4.1 - Introdução. Fontes. Nitrogênio: requerido em grande quantidade pelas plantas. Nitrogênio do solo. Fertilizantes
 4 Fixação do Nitrogênio 1 4.1 - Introdução Nitrogênio: requerido em grande quantidade pelas plantas Nitrogênio do solo Fertilizantes Fontes FBN Nitrogênio: abundante na natureza (Fonte: Hungria et al.,
4 Fixação do Nitrogênio 1 4.1 - Introdução Nitrogênio: requerido em grande quantidade pelas plantas Nitrogênio do solo Fertilizantes Fontes FBN Nitrogênio: abundante na natureza (Fonte: Hungria et al.,
CURSO DE AGRONOMIA FERTILIDADE DO SOLO
 CURSO DE AGRONOMIA FERTILIDADE DO SOLO Prof. Leandro Souza da Silva Prof. Carlos Alberto Ceretta Prof. Danilo R. dos Santos Aula 1 Bases conceituais à fertilidade do solo Fertilidade do solo Solo -Sistema
CURSO DE AGRONOMIA FERTILIDADE DO SOLO Prof. Leandro Souza da Silva Prof. Carlos Alberto Ceretta Prof. Danilo R. dos Santos Aula 1 Bases conceituais à fertilidade do solo Fertilidade do solo Solo -Sistema
Cópia autorizada. II
 II Sugestões de avaliação Ciências 7 o ano Unidade 2 5 Unidade 2 Nome: Data: 1. As cores das flores e o período do dia em que elas abrem são duas adaptações importantes das plantas e que facilitam a sua
II Sugestões de avaliação Ciências 7 o ano Unidade 2 5 Unidade 2 Nome: Data: 1. As cores das flores e o período do dia em que elas abrem são duas adaptações importantes das plantas e que facilitam a sua
PROVA DE CONHECIMENTOS GERAIS Processo Seletivo 2016 MESTRADO. Nome:... Assinatura:...
 Universidade Federal do Amapá Pró-Reitoria de Pesquisa e Pós-Graduação Programa de Pós-Graduação em Biodiversidade Tropical - PPGBIO Mestrado e Doutorado UNIFAP / EMBRAPA-AP / IEPA / CI - BRASIL PROVA
Universidade Federal do Amapá Pró-Reitoria de Pesquisa e Pós-Graduação Programa de Pós-Graduação em Biodiversidade Tropical - PPGBIO Mestrado e Doutorado UNIFAP / EMBRAPA-AP / IEPA / CI - BRASIL PROVA
Ciclos Biogeoquímicos
 Ciclos Biogeoquímicos Matéria orgânica: são os restos dos seres vivos. É composta essencialmente de compostos de carbono. Decompositores: são responsáveis pela degradação da matéria orgânica e favorecem
Ciclos Biogeoquímicos Matéria orgânica: são os restos dos seres vivos. É composta essencialmente de compostos de carbono. Decompositores: são responsáveis pela degradação da matéria orgânica e favorecem
01- Analise a figura abaixo e aponte as capitais dos 3 estados que compõem a Região Sul.
 PROFESSOR: EQUIPE DE GEOGRAFIA BANCO DE QUESTÕES - GEOGRAFIA - 7º ANO - ENSINO FUNDAMENTAL - PARTE 1 ============================================================================================= 01- Analise
PROFESSOR: EQUIPE DE GEOGRAFIA BANCO DE QUESTÕES - GEOGRAFIA - 7º ANO - ENSINO FUNDAMENTAL - PARTE 1 ============================================================================================= 01- Analise
IFRN CICLOS BIOGEOQUÍMICOS
 IFRN CICLOS BIOGEOQUÍMICOS Prof. Hanniel Freitas Ciclos biogeoquímicos Elementos químicos tendem a circular na biosfera. Ciclagem de nutrientes - movimento desses elementos e compostos inorgânicos essenciais
IFRN CICLOS BIOGEOQUÍMICOS Prof. Hanniel Freitas Ciclos biogeoquímicos Elementos químicos tendem a circular na biosfera. Ciclagem de nutrientes - movimento desses elementos e compostos inorgânicos essenciais
Requisitos de Clima e Solo da espécie
 Requisitos de Clima e Solo da espécie Clima O amendoim forrageiro apresenta ampla faixa de adaptação, desenvolvendo-se bem desde o nível do mar até aproximadamente 1.800 m de altitude, em áreas com precipitação
Requisitos de Clima e Solo da espécie Clima O amendoim forrageiro apresenta ampla faixa de adaptação, desenvolvendo-se bem desde o nível do mar até aproximadamente 1.800 m de altitude, em áreas com precipitação
Manejo da adubação nitrogenada na cultura do milho
 Manejo da adubação nitrogenada na cultura do milho Atualmente, pode-se dizer que um dos aspectos mais importantes no manejo da adubação nitrogenada na cultura do milho refere-se à época de aplicação e
Manejo da adubação nitrogenada na cultura do milho Atualmente, pode-se dizer que um dos aspectos mais importantes no manejo da adubação nitrogenada na cultura do milho refere-se à época de aplicação e
Geologia e conservação de solos. Luiz José Cruz Bezerra
 Geologia e conservação de solos Luiz José Cruz Bezerra SOLO É a parte natural e integrada à paisagem que dá suporte às plantas que nele se desenvolvem. Parte mais superficial e fina da crosta terrestre.
Geologia e conservação de solos Luiz José Cruz Bezerra SOLO É a parte natural e integrada à paisagem que dá suporte às plantas que nele se desenvolvem. Parte mais superficial e fina da crosta terrestre.
SOLO. Matéria orgânica. Análise Granulométrica
 SOLO ph Matéria orgânica Análise Granulométrica Disponibilidade dos nutrientes em função do ph Os nutrientes necessários aos vegetais são divididos em duas categorias: Macronutrientes - N, P, K, Ca, Mg,
SOLO ph Matéria orgânica Análise Granulométrica Disponibilidade dos nutrientes em função do ph Os nutrientes necessários aos vegetais são divididos em duas categorias: Macronutrientes - N, P, K, Ca, Mg,
BIOLOGIA - 2 o ANO MÓDULO 27 SUCESSÃO ECOLÓGICA
 BIOLOGIA - 2 o ANO MÓDULO 27 SUCESSÃO ECOLÓGICA Alterações nas condições ambientais Substrato aberto Organismos se estabelecem Novos organismos se estabelecem Estabilização da comunidade e das condições
BIOLOGIA - 2 o ANO MÓDULO 27 SUCESSÃO ECOLÓGICA Alterações nas condições ambientais Substrato aberto Organismos se estabelecem Novos organismos se estabelecem Estabilização da comunidade e das condições
Metas Curriculares. Ensino Básico. Ciências Naturais
 Metas Curriculares Ensino Básico Ciências Naturais 2013 8.º ANO TERRA UM PLANETA COM VIDA Sistema Terra: da célula à biodiversidade 1. Compreender as condições próprias da Terra que a tornam o único planeta
Metas Curriculares Ensino Básico Ciências Naturais 2013 8.º ANO TERRA UM PLANETA COM VIDA Sistema Terra: da célula à biodiversidade 1. Compreender as condições próprias da Terra que a tornam o único planeta
CONCEITOS DE ECOLOGIA. É a história natural científica que se relaciona à sociologia e economia dos animais (Elton,1937)
") CONCEITOS DE ECOLOGIA É a ciência das biocenoses (Clemens, 1916) É a história natural científica que se relaciona à sociologia e economia dos animais (Elton,1937) É a ciência que estuda as leis que regem
CONCEITOS DE ECOLOGIA É a ciência das biocenoses (Clemens, 1916) É a história natural científica que se relaciona à sociologia e economia dos animais (Elton,1937) É a ciência que estuda as leis que regem
17/10/2014 MOLECULAR DOS VEGETAIS. Introdução QUÍMICA DA VIDA. Quais são os elementos químicos encontrados nos Seres Vivos? Elementos Essenciais
 UNIVERSIDADE FEDERAL DE CAMPINA GRANDE CENTRO DE EDUCAÇÃO E SAÚDE CURSO DE BACHARELADO EM FARMÁCIA QUÍMICA DA VIDA Introdução COMPOSIÇÃO MOLECULAR DOS VEGETAIS Quais são os elementos químicos encontrados
UNIVERSIDADE FEDERAL DE CAMPINA GRANDE CENTRO DE EDUCAÇÃO E SAÚDE CURSO DE BACHARELADO EM FARMÁCIA QUÍMICA DA VIDA Introdução COMPOSIÇÃO MOLECULAR DOS VEGETAIS Quais são os elementos químicos encontrados
1. Nitrato de potássio para uma nutrição vegetal eficiente
 Nitrato de potássio é uma fonte única de potássio devido ao seu valor nutricional e a sua contribuição para a sanidade e a produtividade das plantas. O nitrato de potássio possui desejáveis características
Nitrato de potássio é uma fonte única de potássio devido ao seu valor nutricional e a sua contribuição para a sanidade e a produtividade das plantas. O nitrato de potássio possui desejáveis características
Professora Leonilda Brandão da Silva
 COLÉGIO ESTADUAL HELENA KOLODY E.M.P. TERRA BOA - PARANÁ Professora Leonilda Brandão da Silva E-mail: leonildabrandaosilva@gmail.com http://professoraleonilda.wordpress.com/ PROBLEMATIZAÇÃO Como você acha
COLÉGIO ESTADUAL HELENA KOLODY E.M.P. TERRA BOA - PARANÁ Professora Leonilda Brandão da Silva E-mail: leonildabrandaosilva@gmail.com http://professoraleonilda.wordpress.com/ PROBLEMATIZAÇÃO Como você acha
BIE-212: Ecologia Licenciatura em Geociências e Educação Ambiental. Condições e recursos
 BIE-212: Ecologia Licenciatura em Geociências e Educação Ambiental Condições e recursos Programa Introdução Módulo I: Organismos Forrageamento Mecanismos de defesa Seleção sexual e cuidado parental Condições
BIE-212: Ecologia Licenciatura em Geociências e Educação Ambiental Condições e recursos Programa Introdução Módulo I: Organismos Forrageamento Mecanismos de defesa Seleção sexual e cuidado parental Condições
16 EFEITO DA APLICAÇÃO DO FERTILIZANTE FARTURE
 16 EFEITO DA APLICAÇÃO DO FERTILIZANTE FARTURE EM RELAÇÃO AO FORMULADO PADRÃO O objetivo deste trabalho foi avaliar o desempenho do fertilizante Farture (00-12-12) em diferentes dosagens em relação ao
16 EFEITO DA APLICAÇÃO DO FERTILIZANTE FARTURE EM RELAÇÃO AO FORMULADO PADRÃO O objetivo deste trabalho foi avaliar o desempenho do fertilizante Farture (00-12-12) em diferentes dosagens em relação ao
Anais do Seminário de Bolsistas de Pós-Graduação da Embrapa Amazônia Ocidental
 Anais do Seminário de Bolsistas de Pós-Graduação da Embrapa Amazônia Ocidental Anais do Seminário de Bolsistas de Pós-Graduação da Embrapa Amazônia Ocidental Propagação Vegetativa de Piper hispidum Sw.
Anais do Seminário de Bolsistas de Pós-Graduação da Embrapa Amazônia Ocidental Anais do Seminário de Bolsistas de Pós-Graduação da Embrapa Amazônia Ocidental Propagação Vegetativa de Piper hispidum Sw.
UNIVERSIDADE FEDERAL DE GOIÁS UFG - PPGA
 UNIVERSIDADE FEDERAL DE GOIÁS UFG - PPGA CURSO: Pós Graduação Agronomia DISCIPLINA: Classificação e Mapeamento de Solos PROF. RENATA S. MOMOLI Eng. Agrônoma Dra. em Solos e Nutrição de Plantas O QUE É
UNIVERSIDADE FEDERAL DE GOIÁS UFG - PPGA CURSO: Pós Graduação Agronomia DISCIPLINA: Classificação e Mapeamento de Solos PROF. RENATA S. MOMOLI Eng. Agrônoma Dra. em Solos e Nutrição de Plantas O QUE É
Planificação Curricular Anual Ano letivo 2014/2015
 Terra, um planeta com vida Sistema Terra: da célula à biodiversidade 1. Compreender as condições próprias da Terra que a tornam o único planeta com vida conhecida no Sistema Solar 1.1. Identificar a posição
Terra, um planeta com vida Sistema Terra: da célula à biodiversidade 1. Compreender as condições próprias da Terra que a tornam o único planeta com vida conhecida no Sistema Solar 1.1. Identificar a posição
BI63B - ECOSSISTEMAS. Profa. Patrícia C. Lobo Faria
 BI63B - ECOSSISTEMAS Profa. Patrícia C. Lobo Faria pclfaria@uol.com.br http://paginapessoal.utfpr.edu.br/patricialobo Fonte: http://www.portalescolar.net http://exame.abril.com.br/mundo/noticias /fotografa-retrata-urso-polar-no-articoem-pele-e-osso
BI63B - ECOSSISTEMAS Profa. Patrícia C. Lobo Faria pclfaria@uol.com.br http://paginapessoal.utfpr.edu.br/patricialobo Fonte: http://www.portalescolar.net http://exame.abril.com.br/mundo/noticias /fotografa-retrata-urso-polar-no-articoem-pele-e-osso
DIVERSIDADE NA BIOSFERA
 DIVERSIDADE NA BIOSFERA A BIOSFERA - Diversidade Organização biológica Extinção e conservação A CÉLULA - Unidade estrutural e funcional dos seres vivos Constituintes básicos Biologia e Geologia 10º ano
DIVERSIDADE NA BIOSFERA A BIOSFERA - Diversidade Organização biológica Extinção e conservação A CÉLULA - Unidade estrutural e funcional dos seres vivos Constituintes básicos Biologia e Geologia 10º ano
Índice de clorofila em variedades de cana-de-açúcar tardia, sob condições irrigadas e de sequeiro
 Seminário de Iniciação Científica e Tecnológica, 7., 2010, Belo Horizonte Índice de clorofila em variedades de cana-de-açúcar tardia, sob condições irrigadas e de sequeiro Thiago Henrique Carvalho de Souza
Seminário de Iniciação Científica e Tecnológica, 7., 2010, Belo Horizonte Índice de clorofila em variedades de cana-de-açúcar tardia, sob condições irrigadas e de sequeiro Thiago Henrique Carvalho de Souza
Prof. Dr. Francisco Soares Santos Filho (UESPI)
") Prof. Dr. Francisco Soares Santos Filho (UESPI) Visão termodinâmica Alfred J. Lotka trabalhou populações e comunidades em sistemas termodinâmicos. Cada sistema apresenta um conjunto de transformações e
Prof. Dr. Francisco Soares Santos Filho (UESPI) Visão termodinâmica Alfred J. Lotka trabalhou populações e comunidades em sistemas termodinâmicos. Cada sistema apresenta um conjunto de transformações e
MINISTÉRIO DA EDUCAÇÃO UNIVERSIDADE FEDERAL RURAL DE PERNAMBUCO UNIDADE ACADÊMICA DE SERRA TALHADA PROGRAMA PÓS-GRADUAÇÃO EM PRODUÇÃO VEGETAL
 PLANO DE ENSINO I IDENTIFICAÇÃO CURSO: Programa de Pós-Graduação em Produção Vegetal MODALIDADE: Presencial DISCIPLINA: Microbiologia do Solo (PPPV7310) TIPO: ( X ) OBRIGATÓRIA ( X ) OPTATIVA PRÉ-REQUISITO:
PLANO DE ENSINO I IDENTIFICAÇÃO CURSO: Programa de Pós-Graduação em Produção Vegetal MODALIDADE: Presencial DISCIPLINA: Microbiologia do Solo (PPPV7310) TIPO: ( X ) OBRIGATÓRIA ( X ) OPTATIVA PRÉ-REQUISITO:
ECOLOGIA E BIODIVERSIDADE
 ECOLOGIA E BIODIVERSIDADE DIEGO DANTAS AMORIM Contado: diego.dantas@ifmg.edu.br Engenheiro Agrônomo Universidade Vale do Rio Doce UNIVALE Msc. em Agricultura Tropical Universidade Federal do Espírito Santo
ECOLOGIA E BIODIVERSIDADE DIEGO DANTAS AMORIM Contado: diego.dantas@ifmg.edu.br Engenheiro Agrônomo Universidade Vale do Rio Doce UNIVALE Msc. em Agricultura Tropical Universidade Federal do Espírito Santo
Ecossistemas BI63B Profa. Patrícia Lobo Faria
 urso de Graduação em Engenharia Ambiental Ecossistemas BI63B Profa. Patrícia Lobo Faria Ecologia de comunidades: Abordagem geral sobre Distúrbios: Fragmentação de hábitats, efeito de borda, ecologia da
urso de Graduação em Engenharia Ambiental Ecossistemas BI63B Profa. Patrícia Lobo Faria Ecologia de comunidades: Abordagem geral sobre Distúrbios: Fragmentação de hábitats, efeito de borda, ecologia da
UNIDADE IV NUTRIÇÃO MINERAL DE PLANTAS: ABSORÇÃO DE ÍONS 1. INTRODUÇÃO 26/6/2012 FONTES DE NUTRIENTES PARA AS PLANTAS
 UNIDADE IV NUTRIÇÃO MINERAL DE PLANTAS: ABSORÇÃO DE ÍONS 1. INTRODUÇÃO 2. ABSORÇÃO DE ÍONS PELAS RAÍZES 2.1. SELETIVIDADE DE ABSORÇÃO 2.2. O SOLO COMO FORNECEDOR DE NUTRIENTES 2.3. ABSORÇÃO PELAS RAÍZES
UNIDADE IV NUTRIÇÃO MINERAL DE PLANTAS: ABSORÇÃO DE ÍONS 1. INTRODUÇÃO 2. ABSORÇÃO DE ÍONS PELAS RAÍZES 2.1. SELETIVIDADE DE ABSORÇÃO 2.2. O SOLO COMO FORNECEDOR DE NUTRIENTES 2.3. ABSORÇÃO PELAS RAÍZES
Comunidades. Quais são as relações entre diferentes populações num determinado local? Atividades na população
 Quais são as relações entre diferentes populações num determinado local? Atividades na população Comunidade é uma assembleia de populações de espécies que ocorrem juntas no espaço e no tempo. Comunidades
Quais são as relações entre diferentes populações num determinado local? Atividades na população Comunidade é uma assembleia de populações de espécies que ocorrem juntas no espaço e no tempo. Comunidades
Vamos iniciar o estudo da unidade fundamental que constitui todos os organismos vivos: a célula.
 Aula 01 Composição química de uma célula O que é uma célula? Vamos iniciar o estudo da unidade fundamental que constitui todos os organismos vivos: a célula. Toda célula possui a capacidade de crescer,
Aula 01 Composição química de uma célula O que é uma célula? Vamos iniciar o estudo da unidade fundamental que constitui todos os organismos vivos: a célula. Toda célula possui a capacidade de crescer,
Adubação orgânica do pepineiro e produção de feijão-vagem em resposta ao efeito residual em cultivo subsequente
 Seminário de Iniciação Científica e Tecnológica, 9., 2, Belo Horizonte 1 Adubação orgânica do pepineiro e produção de feijão-vagem em resposta ao efeito residual em cultivo subsequente Carlos Henrique
Seminário de Iniciação Científica e Tecnológica, 9., 2, Belo Horizonte 1 Adubação orgânica do pepineiro e produção de feijão-vagem em resposta ao efeito residual em cultivo subsequente Carlos Henrique
Experiências Agroflorestais na Comunidade de Boqueirão. Renato Ribeiro Mendes Eng. Florestal, Msc
 Experiências Agroflorestais na Comunidade de Boqueirão Renato Ribeiro Mendes Eng. Florestal, Msc Localização da área de estudo Dados Climáticos da Região Clima Tropical semi-úmido, com 4 a 5 meses de seca
Experiências Agroflorestais na Comunidade de Boqueirão Renato Ribeiro Mendes Eng. Florestal, Msc Localização da área de estudo Dados Climáticos da Região Clima Tropical semi-úmido, com 4 a 5 meses de seca
EFEITO DOS NÍVEIS DE SALINIDADE DA ÁGUA DE IRRIGAÇÃO NA PRODUÇÃO DO FEIJOEIRO COMUM*
 EFEITO DOS NÍVEIS DE SALINIDADE DA ÁGUA DE IRRIGAÇÃO NA PRODUÇÃO DO FEIJOEIRO COMUM* SILVEIRA, A.L. 1 ; SANTANA, M.J. ; BARRETO, A.C. ; VIEIRA, T.A. 3 * Projeto com apoio da FAPEMIG. 1 Estudante Agronomia
EFEITO DOS NÍVEIS DE SALINIDADE DA ÁGUA DE IRRIGAÇÃO NA PRODUÇÃO DO FEIJOEIRO COMUM* SILVEIRA, A.L. 1 ; SANTANA, M.J. ; BARRETO, A.C. ; VIEIRA, T.A. 3 * Projeto com apoio da FAPEMIG. 1 Estudante Agronomia
AVALIAÇÃO DO USO DO PÓ DE ROCHA NO DESEMPENHO DE DUAS VARIEDADES DE MANDIOCA DE MESA
 AVALIAÇÃO DO USO DO PÓ DE ROCHA NO DESEMPENHO DE DUAS VARIEDADES DE MANDIOCA DE MESA Fernanda Pereira de Souza 1 Eder de Souza Martins 2 Luise Lottici Krahl 3 Eduardo Alano Vieira 4 Josefino de Freitas
AVALIAÇÃO DO USO DO PÓ DE ROCHA NO DESEMPENHO DE DUAS VARIEDADES DE MANDIOCA DE MESA Fernanda Pereira de Souza 1 Eder de Souza Martins 2 Luise Lottici Krahl 3 Eduardo Alano Vieira 4 Josefino de Freitas
-ECOLOGIA APLICADA. Espécies símbolos. Prevenção da Poluição. Conservação de áreas. Preservação da diversidade genética bbbb
 -ECOLOGIA APLICADA * É o estudo dos efeitos causados pelo homem nos sistemas ecológicos, e o consequente manejo desses sistemas e recursos em benefício da sociedade. Espécies símbolos Questões Prevenção
-ECOLOGIA APLICADA * É o estudo dos efeitos causados pelo homem nos sistemas ecológicos, e o consequente manejo desses sistemas e recursos em benefício da sociedade. Espécies símbolos Questões Prevenção
UNIVERSIDADE FEDERAL DO PAMPA TRANSFORMAÇÕES DA MATÉRIA E QUANTIDADES LICENCIATURA EM CIÊNCIAS DA NATUREZA. Uruguaiana, maio de 2016.
 UNIVERSIDADE FEDERAL DO PAMPA TRANSFORMAÇÕES DA MATÉRIA E QUANTIDADES LICENCIATURA EM CIÊNCIAS DA NATUREZA CICLOS BIOGEOQUÍMICOS Uruguaiana, maio de 2016. 1 Na natureza nada se cria, nada se perde, tudo
UNIVERSIDADE FEDERAL DO PAMPA TRANSFORMAÇÕES DA MATÉRIA E QUANTIDADES LICENCIATURA EM CIÊNCIAS DA NATUREZA CICLOS BIOGEOQUÍMICOS Uruguaiana, maio de 2016. 1 Na natureza nada se cria, nada se perde, tudo
ABSORÇÃO FOLIAR. Prof. Josinaldo Lopes Araujo. Plantas cultivadas dividem-se em: Folhas Caule Raízes
 ABSORÇÃO FOLIAR Prof. Josinaldo Lopes Araujo 1 INTRODUÇÃO Plantas cultivadas dividem-se em: Folhas Caule Raízes Cada parte tem uma função definida As folhas absorvem água e nutrientes Porque essa capacidade?
ABSORÇÃO FOLIAR Prof. Josinaldo Lopes Araujo 1 INTRODUÇÃO Plantas cultivadas dividem-se em: Folhas Caule Raízes Cada parte tem uma função definida As folhas absorvem água e nutrientes Porque essa capacidade?
Fatores de Formação do Solo
 Clima Forma ativa e diferencial de atuação na formação do solo; Rochas iguais + condições climáticas diferentes = solos diferentes Rochas diferentes + condições climáticas iguais = solos iguais Sheila
Clima Forma ativa e diferencial de atuação na formação do solo; Rochas iguais + condições climáticas diferentes = solos diferentes Rochas diferentes + condições climáticas iguais = solos iguais Sheila
OS CERRADOS. Entre as plantas do cerrado, podemos citar a sucurpira, o pequi, a copaíba, o angico, a caviúna, jatobá, lobeira e cagaita.
 Os principais biomas brasileiros (biomas terrestres) são: A floresta Amazônica, a mata Atlântica, os campos Cerrados, a Caatinga, o Pampa e o Pantanal. OS CERRADOS Localizados nos estados de Goiás, Tocantins,
Os principais biomas brasileiros (biomas terrestres) são: A floresta Amazônica, a mata Atlântica, os campos Cerrados, a Caatinga, o Pampa e o Pantanal. OS CERRADOS Localizados nos estados de Goiás, Tocantins,
ESTUDO DA VIABILIDADE DE DISPONIBILIZAÇÃO DE POTÁSSIO E FÓSFORO EM SOLOS DE CERRADO COM A UTILIZAÇÃO DO PENERGETIC 4 SAFRAS
 ESTUDO DA VIABILIDADE DE DISPONIBILIZAÇÃO DE POTÁSSIO E FÓSFORO EM SOLOS DE CERRADO COM A UTILIZAÇÃO DO PENERGETIC 4 SAFRAS ANDRÉ L.T. FERNANDES DR. ENGENHARIA DE ÁGUA E SOLO, PROF. UNIUBE E FACULDADES
ESTUDO DA VIABILIDADE DE DISPONIBILIZAÇÃO DE POTÁSSIO E FÓSFORO EM SOLOS DE CERRADO COM A UTILIZAÇÃO DO PENERGETIC 4 SAFRAS ANDRÉ L.T. FERNANDES DR. ENGENHARIA DE ÁGUA E SOLO, PROF. UNIUBE E FACULDADES
ESCOLA BÁSICA 2, 3 LUÍS DE CAMÕES. PROJETO CURRICULAR DA DISCIPLINA DE CIÊNCIAS NATURAIS 5º Ano
 ESCOLA BÁSICA 2, 3 LUÍS DE CAMÕES ANO LETIVO 2014 / 2015 PROJETO CURRICULAR DA DISCIPLINA DE CIÊNCIAS NATURAIS 5º Ano DOMÍNIO: A TERRA UM PLANETA ESPECIAL Onde existe vida? O que é a Biosfera? 1 Compreender
ESCOLA BÁSICA 2, 3 LUÍS DE CAMÕES ANO LETIVO 2014 / 2015 PROJETO CURRICULAR DA DISCIPLINA DE CIÊNCIAS NATURAIS 5º Ano DOMÍNIO: A TERRA UM PLANETA ESPECIAL Onde existe vida? O que é a Biosfera? 1 Compreender
EFEITO DE ADUBAÇÃO NITROGENADA EM MILHO SAFRINHA CULTIVADO EM ESPAÇAMENTO REDUZIDO, EM DOURADOS, MS
 EFEITO DE ADUBAÇÃO NITROGENADA EM MILHO SAFRINHA CULTIVADO EM ESPAÇAMENTO REDUZIDO, EM DOURADOS, MS Carlos Hissao Kurihara (1), Bruno Patrício Tsujigushi (2) Introdução A adubação da cultura do milho safrinha
EFEITO DE ADUBAÇÃO NITROGENADA EM MILHO SAFRINHA CULTIVADO EM ESPAÇAMENTO REDUZIDO, EM DOURADOS, MS Carlos Hissao Kurihara (1), Bruno Patrício Tsujigushi (2) Introdução A adubação da cultura do milho safrinha
ABSORÇÃO DE NUTRIENTES E SÓDIO PELO CAUPI EM SOLOS SALINO-SÓDICOS CORRIGIDOS COM GESSO
 UNIVERSIDADE FEDERAL RURAL DE PERNAMBUCO PÓS-GRADUAÇÃO EM CIÊNCIA DO SOLO DISCIPLINA: SEMINÁRIO II PROFº: MARIO DE ANDRADE LIRA JR. Ciência do Solo ABSORÇÃO DE NUTRIENTES E SÓDIO PELO CAUPI EM SOLOS SALINO-SÓDICOS
UNIVERSIDADE FEDERAL RURAL DE PERNAMBUCO PÓS-GRADUAÇÃO EM CIÊNCIA DO SOLO DISCIPLINA: SEMINÁRIO II PROFº: MARIO DE ANDRADE LIRA JR. Ciência do Solo ABSORÇÃO DE NUTRIENTES E SÓDIO PELO CAUPI EM SOLOS SALINO-SÓDICOS
Água, Sais e Carboidratos
 Água, Sais e Carboidratos A Bioquímica estuda as reações químicas dos organismos vivos e tem revelado inúmeras substancias presentes nas células e em outras que ela participa. A bioquímica estuda as moléculas
Água, Sais e Carboidratos A Bioquímica estuda as reações químicas dos organismos vivos e tem revelado inúmeras substancias presentes nas células e em outras que ela participa. A bioquímica estuda as moléculas
CICLOS BIOGEOQUÍMICOS
 CICLOS BIOGEOQUÍMICOS É o trânsito da matéria entre o meio físico e os seres vivos. Quando os organismos vivos realizam os processos vitais essenciais, eles incorporam moléculas de água, carbono, nitrogênio
CICLOS BIOGEOQUÍMICOS É o trânsito da matéria entre o meio físico e os seres vivos. Quando os organismos vivos realizam os processos vitais essenciais, eles incorporam moléculas de água, carbono, nitrogênio
- Para compreender os solos (conceitos e processos importantes)
") Os Solos - Para compreender os solos (conceitos e processos importantes) - Solo: camada superficial, resultante de rocha decomposta ou triturada. Mistura-se a matéria orgânica. - Intemperismo: ação decompositora
Os Solos - Para compreender os solos (conceitos e processos importantes) - Solo: camada superficial, resultante de rocha decomposta ou triturada. Mistura-se a matéria orgânica. - Intemperismo: ação decompositora
Trufa Negra. Líquen Crustoso
 Líquen Crustoso A II Trufa Negra A I http://treparriscosfieldnotebook.blogspot.co m.br/1992/08/verrucaria-maura.html Trufas negras. Foto: Dream79 / Shutterstock.com Formam crostas que estão fortemente
Líquen Crustoso A II Trufa Negra A I http://treparriscosfieldnotebook.blogspot.co m.br/1992/08/verrucaria-maura.html Trufas negras. Foto: Dream79 / Shutterstock.com Formam crostas que estão fortemente
Ecossistemas DEBIO/UFOP Curso: Engenharia Ambiental
 Ecossistemas DEBIO/UFOP Curso: Engenharia Ambiental Tema 18 Fluxo de Matéria e Energia Aula 1: Produção primária Aula 2: Níveis tróficos Prof. Dr. Roberth Fagundes roberthfagundes@gmail.com www.professor.ufop.br/roberthfagundes
Ecossistemas DEBIO/UFOP Curso: Engenharia Ambiental Tema 18 Fluxo de Matéria e Energia Aula 1: Produção primária Aula 2: Níveis tróficos Prof. Dr. Roberth Fagundes roberthfagundes@gmail.com www.professor.ufop.br/roberthfagundes
Adubação do Milho Safrinha. Aildson Pereira Duarte Instituto Agronômico (IAC), Campinas
, Campinas") Adubação do Milho Safrinha Aildson Pereira Duarte Instituto Agronômico (IAC), Campinas Produtividade, kg/ha Área (milhões ha) AUMENTO DA ÁREA E PRODUTIVIDADE 8000,0 7000,0 6000,0 Total MT 7.012 5000,0
Adubação do Milho Safrinha Aildson Pereira Duarte Instituto Agronômico (IAC), Campinas Produtividade, kg/ha Área (milhões ha) AUMENTO DA ÁREA E PRODUTIVIDADE 8000,0 7000,0 6000,0 Total MT 7.012 5000,0
Diversidade de Fungos Micorrízicos e da Microbiota: Importância na Agricultura
 Universidade de São Paulo Centro de Energia Nuclear na Agricultura Diversidade de Fungos Micorrízicos e da Microbiota: Importância na Agricultura Camila Maistro Patreze & Siu Mui Tsai cpatreze@cena.usp.br
Universidade de São Paulo Centro de Energia Nuclear na Agricultura Diversidade de Fungos Micorrízicos e da Microbiota: Importância na Agricultura Camila Maistro Patreze & Siu Mui Tsai cpatreze@cena.usp.br
Ecologia de Populações e Comunidades
 Ecologia de Populações e Comunidades Profa. Isabel Belloni Schmidt Dept. Ecologia UnB isabels@unb.br Biogeografia O que é Ecologia? O que é Biogeografia Estudo de padrões de distribuição e abundância de
Ecologia de Populações e Comunidades Profa. Isabel Belloni Schmidt Dept. Ecologia UnB isabels@unb.br Biogeografia O que é Ecologia? O que é Biogeografia Estudo de padrões de distribuição e abundância de
Condições edáficas do Nordeste para empreendimentos florestais. Prof. Paulo Rogério Soares de Oliveira UFRN
 Condições edáficas do Nordeste para empreendimentos florestais Prof. Paulo Rogério Soares de Oliveira UFRN Dezembro de 2011 Introdução Edafologia é a ciência que trata da influência dos solos em seres
Condições edáficas do Nordeste para empreendimentos florestais Prof. Paulo Rogério Soares de Oliveira UFRN Dezembro de 2011 Introdução Edafologia é a ciência que trata da influência dos solos em seres
Aula 7 PRODUTIVIDADE DOS ECOSSISTEMAS
 PRODUTIVIDADE DOS ECOSSISTEMAS Aula 7 META Apresentar produtividade primária nos ecossistemas terrestres, os fatores limitantes da produtividade e os padrões de produção primária nos ecossistemas aquáticos.
PRODUTIVIDADE DOS ECOSSISTEMAS Aula 7 META Apresentar produtividade primária nos ecossistemas terrestres, os fatores limitantes da produtividade e os padrões de produção primária nos ecossistemas aquáticos.
Ana P. G. C. Marques Rui S. Oliveira Paula M. L. Castro António O. S. S. Rangel
 ZINC ACCUMULATION AND TRANSLOCATION IN SOLANUM NIGRUM: : EFFECT OF ARBUSCULAR MYCORRHIZAL FUNGI AND MATRIX CONCENTRATION Ana P. G. C. Marques Rui S. Oliveira Paula M. L. Castro António O. S. S. Rangel
ZINC ACCUMULATION AND TRANSLOCATION IN SOLANUM NIGRUM: : EFFECT OF ARBUSCULAR MYCORRHIZAL FUNGI AND MATRIX CONCENTRATION Ana P. G. C. Marques Rui S. Oliveira Paula M. L. Castro António O. S. S. Rangel
CAPÍTULO 3 CORTE DE CIPÓS
 CAPÍTULO 3 CORTE DE CIPÓS Corte de Cipós 37 APRESENTAÇÃO Os cipós são plantas trepadeiras que se desenvolvem sobre os troncos e copas de outras árvores. Os cipós ocorrem em toda a Floresta Amazônica, sendo
CAPÍTULO 3 CORTE DE CIPÓS Corte de Cipós 37 APRESENTAÇÃO Os cipós são plantas trepadeiras que se desenvolvem sobre os troncos e copas de outras árvores. Os cipós ocorrem em toda a Floresta Amazônica, sendo
Quadro 1 - Fatores para conversão de unidades antigas em unidades do Sistema Internacional de Unidades.
 Informação sobre interpretação de Análise de Solo, segundo o Centro de Pesquisa e Desenvolvimento de Solos e Recursos Ambientais, Instituto Agronômico de Campinas. Quadro 1 - Fatores para conversão de
Informação sobre interpretação de Análise de Solo, segundo o Centro de Pesquisa e Desenvolvimento de Solos e Recursos Ambientais, Instituto Agronômico de Campinas. Quadro 1 - Fatores para conversão de
ECOSSISTEMA UNIVERSIDADE ESTADUAL DO CEARÁ CENTRO DE CIÊNCIAS DA SAÚDE CIÊNCIAS BIOLÓGICAS ECOLOGIA
 UNIVERSIDADE ESTADUAL DO CEARÁ CENTRO DE CIÊNCIAS DA SAÚDE CIÊNCIAS BIOLÓGICAS ECOLOGIA ECOSSISTEMA Prof.: Dr. ORIEL HERRERA Monitores: John David & Giuliane Sampaio Fortaleza/CE Versão 2010 ECOSSISTEMA
UNIVERSIDADE ESTADUAL DO CEARÁ CENTRO DE CIÊNCIAS DA SAÚDE CIÊNCIAS BIOLÓGICAS ECOLOGIA ECOSSISTEMA Prof.: Dr. ORIEL HERRERA Monitores: John David & Giuliane Sampaio Fortaleza/CE Versão 2010 ECOSSISTEMA
Biomas no mundo. O conceito de bioma! Os tipos de Bioma
 Biomas no mundo O conceito de bioma! Condições físicas e geográficas, tais como clima, topografia e solo são fatores determinantes no processo de seleção natural que leva a diferenciação das espécies.
Biomas no mundo O conceito de bioma! Condições físicas e geográficas, tais como clima, topografia e solo são fatores determinantes no processo de seleção natural que leva a diferenciação das espécies.
Biomassa de Banana Verde Polpa - BBVP
 Biomassa de Banana Verde Polpa - BBVP INFORMAÇÕES NUTRICIONAIS Porção de 100g (1/2 copo) Quantidade por porção g %VD(*) Valor Energético (kcal) 91 4,55 Carboidratos 21,4 7,13 Proteínas 2,1 2,80 Gorduras
Biomassa de Banana Verde Polpa - BBVP INFORMAÇÕES NUTRICIONAIS Porção de 100g (1/2 copo) Quantidade por porção g %VD(*) Valor Energético (kcal) 91 4,55 Carboidratos 21,4 7,13 Proteínas 2,1 2,80 Gorduras
Características gerais
 Características gerais Organismos eucariontes, unicelulares ou pluricelulares; Podem ser de vida livre ou não; Podem ser encontrados nos mais variados ambientes, preferencialmente em lugares úmidos; Característica
Características gerais Organismos eucariontes, unicelulares ou pluricelulares; Podem ser de vida livre ou não; Podem ser encontrados nos mais variados ambientes, preferencialmente em lugares úmidos; Característica
Agrupamento Escolas José Belchior Viegas - Escola E.B. 2,3 Poeta Bernardo de Passos Ciências Naturais Planificação anual 5ºAno Ano letivo:
 Agrupamento Escolas José Belchior Viegas - Escola E.B. 2,3 Poeta Bernardo de Passos Ciências Naturais Planificação anual 5ºAno Ano letivo: 2015-16 Período Conteúdos INTRODUÇÃO Onde existe vida? Primeira
Agrupamento Escolas José Belchior Viegas - Escola E.B. 2,3 Poeta Bernardo de Passos Ciências Naturais Planificação anual 5ºAno Ano letivo: 2015-16 Período Conteúdos INTRODUÇÃO Onde existe vida? Primeira
Troca de materiais entre os componentes bióticos e abióticos dos ecossistemas.
 Troca de materiais entre os componentes bióticos e abióticos dos ecossistemas. CICLO do FÓSFORO CICLO SEDIMENTAR APATITA Ca 3 (PO 4 ) 2 erosão de rochas fosfatadas CICLO RÁPIDO CICLO LENTO PICO DO FÓSFORO
Troca de materiais entre os componentes bióticos e abióticos dos ecossistemas. CICLO do FÓSFORO CICLO SEDIMENTAR APATITA Ca 3 (PO 4 ) 2 erosão de rochas fosfatadas CICLO RÁPIDO CICLO LENTO PICO DO FÓSFORO
18 PRODUTIVIDADE DA SOJA EM FUNÇÃO DA
 18 PRODUTIVIDADE DA SOJA EM FUNÇÃO DA APLICAÇÃO DE MACRONUTRIENTES EM PÓS- EMERGÊNCIA DA CULTURA O objetivo neste trabalho foi avaliar a aplicação de macronutrientes de diversas fontes e épocas de aplicação
18 PRODUTIVIDADE DA SOJA EM FUNÇÃO DA APLICAÇÃO DE MACRONUTRIENTES EM PÓS- EMERGÊNCIA DA CULTURA O objetivo neste trabalho foi avaliar a aplicação de macronutrientes de diversas fontes e épocas de aplicação
UNESP Universidade Estadual Paulista Campus Experimental de Dracena Fazuldade de Zootecnia. Curso: Zootecnia Professor: Dr. Reges Heinrichs 2010
 FÓSFORO UNESP Universidade Estadual Paulista Campus Experimental de Dracena Fazuldade de Zootecnia Curso: Zootecnia Professor: Dr. Reges Heinrichs 2010 FÓSFORO INTRODUÇÃO FORMAS NO SOLO DISPONIBILIDADE
FÓSFORO UNESP Universidade Estadual Paulista Campus Experimental de Dracena Fazuldade de Zootecnia Curso: Zootecnia Professor: Dr. Reges Heinrichs 2010 FÓSFORO INTRODUÇÃO FORMAS NO SOLO DISPONIBILIDADE
Ecossistemas I. Umberto Kubota Laboratório de Interações Inseto-Planta Dep. Zoologia IB Unicamp. Produtividade secundária
 Ecossistemas I Umberto Kubota ukubota@gmail.com Laboratório de Interações Inseto-Planta Dep. Zoologia IB Unicamp Ecossistemas I Conceitos básicos Fluxo de energia Produtividade primária Ecossistema: Conceitos
Ecossistemas I Umberto Kubota ukubota@gmail.com Laboratório de Interações Inseto-Planta Dep. Zoologia IB Unicamp Ecossistemas I Conceitos básicos Fluxo de energia Produtividade primária Ecossistema: Conceitos
A qualidade de mudas clonais de Eucalyptus urophylla x E. grandis impacta o aproveitamento final de mudas, a sobrevivência e o crescimento inicial
 Anais da 50ª Reunião Técnico-Científica do Programa Cooperativo sobre Silvicultura e Manejo - Dias 12 e 13 de novembro de 2014 33 A qualidade de mudas clonais de Eucalyptus urophylla x E. grandis impacta
Anais da 50ª Reunião Técnico-Científica do Programa Cooperativo sobre Silvicultura e Manejo - Dias 12 e 13 de novembro de 2014 33 A qualidade de mudas clonais de Eucalyptus urophylla x E. grandis impacta
ECOSSISTEMA. inseparavelmente ligados. Organismos vivos + Ambiente (abiótico) interagem entre si
 interagem entre si") ECOSSISTEMA inseparavelmente ligados Organismos vivos + Ambiente (abiótico) interagem entre si Ciclos de Diversidade Estrutura Materiais Biótica Trófica Troca de materiais vivos e não vivos Componente
ECOSSISTEMA inseparavelmente ligados Organismos vivos + Ambiente (abiótico) interagem entre si Ciclos de Diversidade Estrutura Materiais Biótica Trófica Troca de materiais vivos e não vivos Componente
Líquens. LÍQUENS -Introdução
 Líquens LÍQUENS -Introdução Simbiose viver junto é uma associação íntima e duradora entre organismos de espécies diferentes Mutualismo associação benéfica para ambos os organismos Líquen associação simbiótica
Líquens LÍQUENS -Introdução Simbiose viver junto é uma associação íntima e duradora entre organismos de espécies diferentes Mutualismo associação benéfica para ambos os organismos Líquen associação simbiótica