CAMARÕES (ATYIDAE E PALAEMONIDAE) DA REDE FLUVIAL DO ESTADO DO RIO DE JANEIRO: SISTEMÁTICA E DISTRIBUIÇÃO
|
|
|
- André Monsanto de Caminha
- 5 Há anos
- Visualizações:
Transcrição
1 11 Kátia Maria Leal da Fonseca CAMARÕES (ATYIDAE E PALAEMONIDAE) DA REDE FLUVIAL DO ESTADO DO RIO DE JANEIRO: SISTEMÁTICA E DISTRIBUIÇÃO Banca Examinadora: Prof. Dr Idalina Maria Brasil Li.ma (Presidente da Banca) Prof. Dr. Jorge Luiz Nessimian Prof. Dr. Gustavo Augusto Schimidt Melo Rio de Janeiro,14 de Dezembro de 1995.
2 111 "Quando sabemos qual é o nosso propósito, o trabalho da Alma se realiza da melhor maneira possivel através do nosso corpo. Um propósito claro elimina todas as dúvidas, pois imediatamente identificamos aquilo que nos conduz à nossa meta ou nos desvia dela. A energia em nossas vidas é imensa quando uma clareza de propósito está sempre presente. " (tirado do livro Meditando com os Anjos de Sônia Café)
3 lv Trabalho realizado no Departamento de Zoologia, Instituto de Biologia e no Departamento de Invertebrados, Museu Nacional - Universidade Federal do Rio de Janeiro. 1! Orientadora: Profa. Dt:'1 Maria Margarida Gomes Corrêa Universidade Federal do Rio de Janeiro Museu Nacional
4 V FICHA CATALOGRÁFICA FONSECA, Kátia Maria Leal Camarões de água doce (Atyidae e Palaemonidae) da rede fluvial do Estado do Rio de Janeiro: sistemática e distribuição. Rio de Janeiro. UFRJ, Museu Nacional, xi, 85 f. Tese: Mestre em Ciências Biológicas (Zoologia) 1. Atyidae e Palaemonidae 2. Sistemática 3. Distribuição 4. Rio de Janeiro I. Universidade Federal do Rio de Janeiro - Museu Nacional II. Teses
5 VI AGRADECIMENTOS À Professora Doutora Maria Margarida Gomes Corrêa, do Museu Nacional / UFRJ, pelo incentivo e orientação durante o desenvolvimento deste trabalho. À Professora Maria Cristina Ostrovski de Matos, do Departamento de Zoologia / I.B. / UFRJ, pelo amparo técnico e importantes sugestões dadas durante conversas informais, as quais enriqueceram este trabalho. À Professora Doutora Érica Caramaschi, do Departamento de Ecologia/ IB / UFRJ, por ter cedido parte do material estudado. Ao Professor Doutor Gustavo Augusto Schmidt de Melo, do Museu de Zoologia da Universidade de São Paulo, pela atenção dada e por permitir a consulta ao material de interesse para este trabalho, pertencente à coleção do MZUSP. Ao Professor Doutor Jorge Soares Marques, do Departamento de Geografia/ UFRJ, pela atenção a mim dispensada e pelas importantes informações e sugestões dadas, que enriqueceram este trabalho. Ao Professor Doutor Sérgio Luiz Costa Bonecker, do Departamento de Zoologia/ IB / UFRJ, pela utilização de seu equipamento de informática. À Edelti Faria Albertoni, do Laboratório de Limnologia, Departamento de Ecologia / UFRJ, por ter doado alguns exemplares e por sua
6 Vll contribuição com o fornecimentos de dados importantes que ainda faltavam neste trabalho. À grande arruga Margaret M ª. de Oliveira Corrêa, p01s sem seu incentivo e ajuda talvez não houvesse realizado este trabalho. À também grande amiga Geórgia Corrêa Atella pelo ap010 dado sempre que necessário. Ao amigo Flávio A. Bockmann por toda a sua boa vontade em ajudar, e também pelo incentivo dado. A todos os meus amigos que, com seu apoio e compreensão tomaram possível a realização deste trabalho. À Coordenação do Curso de Pós-Graduação em Zoologia / Museu Nacional e do Departamento de Zoologia / Instituto de Biologia - Universidade Federal do Rio de Janeiro, que tomaram viável a realização deste trabalho em todas as suas etapas. Ao CNPq (Conselho Nacional de Desenvolvimento Técnico e Científico) pelo amparo financeiro cedido sob a forma de bolsa de Mestrado. E, finalmente, à minha mãe, pelo seu incansável apoio e compreensão ao longo de toda a minha vida e sem os quais talvez não houvesse chegado à metade desta caminhada.
7 Vlll RESUMO Esta dissertação compreende um estudo da sistemática e da distribuição geográfica dos camarões de água doce (Atyidae e Palaemonidae) existentes na rede fluvial do Estado do Rio de Janeiro. O presente estudo vem contribuir para o conhecimento de alguns aspectos ambientais ( salinidade, temperatura, declividade e oxigênio) os quais influenciam a distribuição destes arumais. São considerados, também, os mecanismos de dispersão por eles utilizados. Este trabalho é constituído de uma introdução, uma parte sistemática, discussão, conclusões, referências bibliográficas, três quadros, cinco mapas e dez estampas das espécies estudadas. São apresentadas chaves de identificação para famílias e espécies, diagnoses das famílias e subfamílias, caracterização das espécies, tamanho, localidade tipo, distribuição geográfica, material examinado e observações. É registrada a ocorrência de uma espécie ainda não assinalada para a região, questionada a validade de uma espécie; e também são referidos fatores que influenciam a distribuição das espécies. As espécies são distribuídas de acordo com as unidades hidrográficas delimitadas pela SERLA (Superintendência Estadual de Rios e Lagos) em 1985.
8 IX SHRIMPS (ATYIDAE AND PALAEMONIDAE) FROM THE RIO DE JANEIRO FLUVIAL SYSTEM: SYSTEMATICS AND DISTRIBUTION. ABSTRACT Systematics and geographic distribution of freshwater shrimps (Atyidae and Palaemonidae) from the Rio de Janeiro fluvial system were studied. Some environmental factors (salinity, temperature, declivity and oxygen) influence upon species distribution. This dissertation contains an introduction, systematic part, discussion, conclusions, bibliography, three squares, five maps and ten plates corresponding to the species studied. Keys for classification of families and species, diagnosis of families and subfamilies are shown. Characterization, size, type locality, geographic distribution, material examined and notes for each species are mentioned. The occurrence of one species not yet recorded from the region is mentioned, the validity of one species is discussed, and reported factors which influence upon the species distribution. The species are distributed according to the hydrographic basins delimitated for SERLA (Superintendência Estadual de Rios e Lagos) in 1985.
9 X ÍNDICE Agradecimentos Resumo Abstract 1. Introdução Objetivos H. t'. '... 1s onco d as espec1es encon tr a d as Distribuição das espécies pelo continente Fatores ainbientais... 5 Salinidade... 5 Temperatura... 6 O xigemo." Declividade... 7 Alguns mecanismos de dispersão Material e Métodos Coleções estudadas No Cainpo No laboratório Parte Sistemática Chave para famílias Família Atyidae De Haan, Chave para as espécies da Família Atyidae Gênero A tya Leach,
10 XI 1) Atya gabonensis Giebel, ) A. scabra (Leach, 1815) Gênero Potimirim Holthuis, ) Potimirim glabra (Kingsley, 1878) ) P. potimirim (Müller, 1881) Família Palaemonidae Rafinesque, Subfamília Palaemoninae Dana, Chave para as espécies da Família Palaemonidae Gênero Palaemon Weber, ) Pa/aemon (Palaemon) panda/iformis (Stimpson, 1871) Gênero Macrobrachium Bate, ) Macrobrachium acanthurus (Wiegmann, 1836) ) M carcinus (Linnaeus, 1758) ) M olfersii (Wiegmann, 1836) ) M potiuna (Müller, 1880) ) M iheringi (Ortmann, 1897) Discussão Sistemática Distribuição Conclusões Referências bibliográficas Mapas Estampas
11 1 1. INTRODUÇÃO Segundo RODRIGUEZ (1981) as famílias Atyidae e Palaemonidae estão representadas por camarões de hábitos bentônicos. Os Atyidae estão quase completamente confinados ao ambiente de água doce, estando representados no Estado do Rio de Janeiro pelos gêneros Atya Leach, 1816 e Potimirim Holthuis, 1954, sendo o primeiro pantropical e o segundo endêmico da região neotropical. A família Palaemonidae, de ampla distribuição no mundo, inclui os camarões de rio do gênero Macrobrachium Bate, 1868, pantropicais. ESCOBAR (1979) considera os Palaemonidae como animais não muito exigentes quanto à dieta alimentar, alimentando-se de peixes, crustáceos, larvas e adultos de insetos, plantas terrestres e aquáticas, folhas, sementes e algas, e os Atyidae também com alimentação detritivora. Segundo HOBBS & HART (1982), os Atyidae são mais ativos à noite do que durante o dia e COELHO (1963) considerou o mesmo quanto aos Palaemonidae. Das quatro subfamílias pertencentes à família Palaemonidae (Palaemoninae, Pontoniinae, Euryrhynchinae e Typhlocaridinae ), apenas uma, a subfamília Palaemoninae está representada na rede fluvial do Rio de Janeiro OBJETIVOS: O presente trabalho tem como objetivo o levantamento da fauna de camarões de água doce (Palaemonidae e Atyidae) do Estado do Rio de Janeiro,
12 compreendendo um estudo da sistemática, aspectos relacionados à distribuição geográfica, das espécies nas bacias hidrográficas do Estado do Rio de Janeiro IDSTÓRICO DAS ESPÉCIES ENCONTRADAS: O gênero Macrobrachium é o mais amplamente distribuído. O pnmerro registro para o Brasil de um espécimen do gênero foi feito por MARCGRAF (1648), que chamou de "Potipema", a espécie que mais tarde foi identificada como Macrobrachium carcinus (Linnaeus, 1758). Em 1880, MÜLLER encontrou um novo camarão de água doce do Brasil, o qual chamou de Pa/aemon potiuna, que mais tarde foi incluído no gênero Macrobrachium. ORTMANN, em 1897 ampliou o número de espécies de camarões de água doce para o Brasil, com o registro de duas ocorrências novas, Pa/aemon acanthurus Wiegmann, 1836 e Palaemon olfersii Wiegmann, 1836 e a descrição da nova espécie Palaemon iheringi, todas sendo, posteriormente, incorporadas ao gênero Macrobrachium. No mesmo ano IHERING, listou os camarões de água doce do Brasil, adicionando algumas informações sobre as localidades onde foram encontradas. O trabalho de MOREIRA (1901) sobre os crustáceos do Brasil, também foi uma grande contribuição para o conhecimento dos camarões de água doce.
13 3 Mais uma importante contribuição para o melhor conhecimento da fauna de camarões de água doce do Brasil, foi dada por SA W A YA em 1946, o qual incluiu uma chave de identificação para machos e fêmeas. Outros importantes trabalhos trataram da família Palaemonidae, como a revisão feita por HOLTHUIS em 1952 sobre a subfamília Palaemoninae, e as publicações de WILLIAMS (1965, 1984) e CHACE & HOBBS (1969) que contêm importantes informações. GOMES-CORRÊA (1977) no trabalho intitulado Palaemonídeos do Brasil, tratou de 28 espécies, incluindo chaves de identificação para as espécies brasileiras. A família Atyidae, mostra-se representada no Estado Rio de Janeiro por dois gêneros (Atya e Potimirim) e quatro espécies. O primeiro registro que temos de um espécimen do gênero Atya no Brasil, foi encontrado em MARCGRAF (1648) o qual chamou "Guaricuru". Em 1962, LEMOS DE CASTRO identificou a gravura de Marcgraf como Atya scabra Leach, LEACH (1815) primeiramente descreveu Atys scaber de local desconhecido. Em 1816 corrigiu o nome para Atya scabra após ter descoberto que o gênero Atys pertencia a um molusco gastrópodo. O gênero então criado por LEACH, posteriormente tomou-se gênero tipo da família Atyidae. Em 1864 A. MILNE-EDW ARDS publicou a primeira revisão do grupo em seu trabalho intitulado "Revision des Crustacés Macroures de la Familie des Atyoidées ". MÜLLER (1881) descreveu uma espécie do Brasil que chamou de Atyoida potimirim, a qual foi mais tarde sinonimizada para Ortmannia potimirim
14 4 por RATHBUN, Em 1954, HOLTHUIS considerou sinônimos os gêneros Ortmannia Rathbun, 1901 e Atya Leach, 1816, baseado em estudos feitos por EDMONSON (1929) e RADIR (1930) (apud HOLTHUIS, 1954) os quais descobriram que a espécie tipo do gênero Ortmannia, O. henshawi Rathbun, 1902, era idêntica à Atya bisulcata Sharp, RATHBUN em 1901 observou que a espécie tipo do gênero Atyoida, A. bisulcata (Randall, 1839), era idêntica à Atya bisulcata. HOLTHUIS (1954), baseado no que havia sido observado por RATHBUN, considerou Atyoida como sinônimo de Atya e propôs o nome Potimirim para o gênero das outras espécies de Atyoida. Em 1904, BOUVIER publicou uma revisão dos Atyidae do Museu de História N aturai de Paris, e em 1925 descreveu as variações e a distribuição geográfica desta família. Em seu trabalho sobre os Atyidae do México, VILLALOBOS (1959) descreveu uma espécie nova para o Brasil, Potimirim brasiliana, baseado na análise de um material proveniente do rio Ariró, Angra dos Reis, Estado do Rio de Janeiro. Em 1963, SMALLEY sugeriu que P. brasiliana fosse colocada como sinônima de P. glabra (KINGSLEY, 1878), devido à sua grande semelhança com esta espécie. CHACE & HOBBS (1969) incluiram P. brasiliana, pela primeira vez, na lista de sinonímia de P. glabra. Outros autores têm citado representantes desta família em seus trabalhos, HART (1961); CHACE (1972); ABELE, (1972); HUNTE (1978); RODRIGUEZ (1980, 1981); VILLALOBOS (1982); ABELE & KIM (1986, 1989). Em 1980 HOBBS citou a ocorrência de Atya gabonensis Giebel, 1875 pela primeira vez no Brasil, mais especificamente no Estado do Piauí.
15 5 Uma publicação de HOBBS & HART (1982), representou grande contribuição para o estudo do gênero Atya, com chaves de identificação para todas as espécies do gênero DISTRIBUIÇÃO DAS ESPÉCIES PELO CONTINENTE: Os padrões de distribuição geográfica das espécies de camarões de água doce são influenciados por alguns fatores ambientais e também por mecanismos de dispersão utilizados por estas espécies FATORES AMBIENTAIS: SALINIDADE Segundo HUNTE (1978) os camarões de água doce, assim como os camarões marinhos, tiveram sua origem no mar, migrando do ambiente marinho para o de água doce, no curso de sua evolução. RODRIGUEZ (1981) observou que alguns destes camarões têm a necessidade de uma determinada concentração de cloretos no seu desenvolvimento larvar, fato este que justifica sua dependência do estuário. A partir destas observações Rodriguez (op. cit) classificou estes camarões em espécies litorâneas e espécies continentais. Alguns outros autores têm
16 realizado trabalhos nos quais citam hábitos migratórios dos camarões de ambas as famílias (Palaemonidae e Atyidae ). 6 COELHO & RAMOS-PORTO (1985) seguem a classificação de RODRIGUEZ (1981) definindo-a da seguinte forma: espécies litorâneas - São encontradas nas bacias costeiras e no curso inferior dos grandes rios, pois suas fêmeas precisam migrar para o estuário no período de desova. espécies continentais - São confinadas aos habitats de água doce. Segundo VILLALOBOS (1982), estas espécies apresentam-se perfeitamente adaptadas aos hábitos dulciaquícolas pelo fato de produzirem ovos grandes, os quais vão eclodir em um estágio mais avançado, como resultado do encurtamento dos estágios poslarvares dependentes do estuário. TEMPERA TURA Segundo CHRISTOFOLETTI (1981), a energia potencial dos rios é convertida, no fluxo, em energia cinética que, por sua vez, é grandemente dissipada em calor e fricção, sendo grande parte da energia de um rio consumida em calor. Dessa forma depois de uma cachoeira ou trecho encachoeirado, a água do rio estará mais quente do que antes de passar por esse trecho. Além disso, as águas que vêm de áreas mais elevadas são mais frias, e para chegarem às áreas mais baixas descem por gradientes elevados. Nas áreas de elevado gradiente 1, a velocidade das águas é maior, portanto o tempo de insolação é menor; em áreas de velocidade menor, maior é o tempo de insolação acarretando um aumento de temperatura da água. 1 O gradiente está relacionado à declividade do terreno. Quanto maior a inclinação, maior será o gradiente.
17 7 OXIGÊNIO A superficie da água tem uma área de contato com o ar. Com a turbulência das águas essa área aumenta e, inclusive, águas profundas chegam à superficie. Portanto quanto maior a turbulência, maior a oxigenação. Trechos com muitos blocos ou muitos afloramentos rochosos, provocam turbulência o que ajuda na oxigenação, apesar da redução de velocidade do fluxo (J. S. MARQUES, com. pes.). De acordo com HUNTE (1978) a quantidade de oxigênio dissolvido na água está diretamente relacionada ao gradiente de cada trecho do rio ( quanto maior o gradiente, maior a velocidade de fluxo). Trechos de rio com gradiente alto têm um alto nível de oxigênio, o que provavelmente ocorre devido à grande turbulência e mistura de suas águas. De acordo com experimentos realizados pelo mesmo autor, espécies que vivem em trechos de rios de baixo gradiente, geralmente são mais tolerantes a baixos níveis de oxigênio. DECLIVIDADE A velocidade do fluxo de águas de um rio é influenciada pela declividade do terreno, presença de obstáculos e espraiamento de água, ou seja, quanto maior a declividade do terreno por onde passa o rio maior será a velocidade de suas águas. A velocidade é medida num ponto ou num trecho, ou seja ao longo de todo o rio o fluxo apresenta vários valores de velocidade. A declividade também influenciará no tipo de leito do rio, pois quanto maior for o declive do terreno maior será a velocidade de corrente provocando uma maior erosão das margens (lateral) e do leito (vertical). Essa maior erosão (vertical e lateral) será responsável pela obtenção de sedimentos, que depositar-se-ão mais adiante, onde houver condições favoráveis, comumente no baixo curso dos rios, predominando quase sempre
18 8 sedimentos arenosos ou lamosos no fundo de seu leito. De acordo com estes fatores as espécies continentais e litorâneas estarão relacionadas a fundo de pedras, no primeiro caso, e fundo de areia e lodo, no segundo ALGUNS MECANISMOS DE DISPERSÃO Os animais dulciaquícolas, têm o seu potencial de dispersão limitado, pelo fato de habitarem massas d'água isoladas. A retenção do estágio planctônico marinho vem resolver esse problema, mantendo seu potencial de dispersão. Alguns outros mecanismos são utilizados como migrações rio acima, transpondo pequenas cachoeiras e deslocamentos fora da água.
19 9 2. MATERIAL E MÉTODOS 2.1. COLEÇÕES ESTUDADAS: Uma parte do material examinado está depositada na coleção Carcinológica do Departamento de Zoologia - Instituto de Biologia da Universidade Federal do Rio de Janeiro e a outra é pertencente à coleção do Setor de Carcinologia - Departamento de Invertebrados - Museu Nacional - UFRJ. Este material inclui espécimens coletados pela equipe do Setor de Ictiologia do Departamento de Vertebrados - Museu Nacional do Rio de Janeiro e coordenada pela Prof! Dr Érica Caramaschi do Departamento de Ecologia - Instituto de Biologia - UFRJ. Também fazem parte da referida coleção, espécimens doados pela equipe do laboratório de Herpetologia do Departamento de Zoologia - Instituto de Biologia - UFRJ. Alguns exemplares incluidos neste trabalho são pertencentes à coleção carcinológica do Museu de Zoologia da Universidade de São Paulo (MZUSP) NO CAMPO: EQUIPAMENTO DE COLETA: Peneira de pedreiro.- Com malha de 3 mm. Utilizada na captura de indivíduos que ficam junto à vegetação marginal. Armadilhas. - Improvisadas com garrafas plásticas ( de refrigerante) com capacidade de 2 litros. Para montá-la basta medir 15 cm do comprimento que
20 10 inclui a boca da garrafa, fazer wn corte transversal, inverter esta parte e em seguida encaixá-la no corpo da garrafa (Fig. 1). A entrada não deve ficar muito grande, para que não entre caranguejos, pois isso impediria a entrada de camarões. Como isca, usa-se carne moída. Também foram colocadas algwnas pedras no interior da armadilha, para que ela não flutuasse. Utilizada na captura de animais que ficam escondidos sob pedras e troncos de árvores. O tempo de espera mínimo, foi de 1 hora. Após a coleta os exemplares foram anestesiados com mentol e depois fixados com álcool à 70% NO LABORA TÓRIO: No laboratório os espécimens foram conservados também em álcool à 70%, e analisados com microscópio estereoscópio OL YMPUS com awnento de até 400 vezes. Foram elaboradas chaves de identificação baseadas em RODRIGUEZ (1980) e HOBBS & HART (1982) para Atyidae e RODRIGUEZ (1980) e HOLTHUIS (1952) para Palaemonidae, adaptadas para as espécies da região em estudo. Os desenhos das peças com as características importantes para a identificação das espécies foram feitas com auxílio de microscópio estereoscópio com câmara clara JENA com awnento de até 100 vezes. A mensuração dos exemplares foi feita com o auxílio de paquímetro, medindo os exemplares do ápice do rostro à extremidade posterior do telso ( comprimento total).
21 11 A caracterização das espécies, foi feita baseando-se em espécimens machos, evidenciando-se os detalhes morfológicos de maior importância para a identificação das espécies. A distribuição geográfica foi baseada em informações adquiridas em literatura, e no material estudado. As espécies encontradas foram distribuídas em unidades hidrográficas delimitadas através de mapeamento feito pela SERLA (Superintendência Estadual de Rios e Lagos) em 1985 e estão expressas no quadro II e nos mapas 1 a 5. O item "Observações" consiste em informações colhidas em literatura, acerca do habitat e problemas sistemáticos. A terminologia utilizada neste trabalho é a mesma adotada por quase todos os especialistas em Decapoda, inclusive RODRIGUEZ, 1980 e HOBBS & HART, 1982, e pode ser consultada nas figuras esquemáticas 2 e 3. Para cada espécie é dada uma lista de material examinado com suas localidades organizadas no sentido norte-sul. Cada procedência é associada a uma abreviatura e um número, que indicam a coleção e o número de registro dos espécimens ou lotes nas coleções a que pertencem. ABREVIA TURAS: DZUFRJ - Departamento de Zoologia da Universidade Federal do Rio de Janeiro. MNRJ - Museu Nacional, Rio de Janeiro, UFRJ. MZUSP - Museu de Zoologia, Universidade de São Paulo.
22 12 15 cm 2.5 cm [ 10cm A B Figura 1: Esquema de montagem de uma annadilha. A Garrafa de refrigerante com capacidade de 2 litros (linha de corte pontilhada), B. Annadilha pronta.
23 13 r--abdome ,-,-- Carapaça Dra.resis Fi gu ra 2: Esquema de um Caridea : ba.- base; ea.- espinho antenal; eh.- espinho branquiostegal; ef.- escafocerito; eh.- espinho hepático; ep.- espinho pterigostômico; ex.- coxa; C.- carpo; D.- dátilo; 1.- ísquio; M.- mero; P.- própode. Fonte: Rodriguez, 1980.

; C.")
24 14 A --a.m. --a.i. B e Figura 3: A Vista lateral do terceiro pereópode do macho adulto de Atya (Hobbs & Hart, 1982); B. Vista lateral do segundo pereópode de Atya (Hobbs & Hart, 1982); C. Segundo pleópode de um macho de Macrobrachium (Gomes-Corrêa, 1977). a.m., apêndice masculino; a.i., apêndice interno.
25 15 3. PARTE SISTEMÁTICA A terminologia e a classificação adotadas seguem basicamente ABELE & BOWMAN, 1982, ABELE, 1992 e BRUSCA & BRUSCA, Subfilo CRUSTACEA Pennant, Classe MALACOSTRACA Latreille, Subclasse EUMALACOSTRACA Grobben, Superordem EUCARIDA Calman, Ordem DECAPODA Latreille, Subordem PLEOCYEMA TA Burkenroad, Infraordem CARIDEA Dana, Superfamília ATYOIDEA De Haan, Família ATYIDAE De Haan, Superfamília PALAEMONOIDEA Rafinesque, Família PALAEMONIDAE Rafinesque, Subfamília PALAEMONINAE Dana, 1852
26 CHAVE PARA AS FAMÍLIAS REPRESENTADAS NO ESTADO DO RIO DE JANEIRO Dedos das quelas do 2 primeiros pares de pereópodes com tufos de longos pêlos tenninais. Rostro desarmado dorsalmente... Atyidae. 2. Dedos das quelas do 2 primeiros pares de pereópodes sem tufos de longos pêlos tenninais. Rostro armado dorsalmente... Palaemonidae. FAMÍLIA ATYIDAE De Haan, Atyidae De Haan, Kingsley, 1878:91 - Chace & Hobbs, 1969:56 - Chace, 1972: 13 - Rodriguez, 1980: : Villalobos, 1982:216. Diagnose.- Rostro curto. Carapaça lisa dorsalmente e somente com espinho antenal e pterigostômico. Dedos dos dois primeiros pares de pereópodes, tenninam em pincéis de longos pêlos (exceto em Xiphocaris von Martens, 1872). Mandíbulas não apresentam palpos. Carpo do segundo pereópode inteiro. Exópodes podem aparecer ou não em todos os pereópodes. Telso alargado e com extremidade posterior arredondada.
27 CHAVE PARA ESPÉCIES DA FAMÍLIA ATYIDAE DO ESTADO DO RIO DE JANEIRO Carpo do segundo par de pereópodes mais largo do que longo. Rostro com margem ventral sem dentes... 2 Carpo do segundo par de pereópodes muito mais longo do que largo. Rostro com margem ventral provida de dentes Carapaça fortemente esculpida. Superficie flexora do própode do terceiro par de pereópodes com tubérculos não arranjados em fileiras longitudinais; dátilo do terceiro par de pereópodes com um grupo de tubérculos na sua superficie flexora sem formar fileiras... Atya gabonensis (est. 1). Carapaça fracamente esculpida. Superficie flexora do própode do terceiro par de pereópodes com fileiras longitudinais de tubérculos; dátilo do terceiro par de pereópodes com tubérculos arranjados em fileira na sua superficie flexora... Atya scabra (est. II). 3. Rostro curvando-se para baixo; apêndice masculino sem espinhos na depressão entre o 2 º e o 3 lobos; com epipodito nos 4 primeiros pares de pereópodes... Potimirim glabra ( est. III). Rostro quase reto; apêndice masculino com espinhos na depressão entre o 2 º e o 3 º lobos; com epipodito nos 3 pnmerros pares de pereópodes... Potimirim potimirim ( est. IV).
28 18 Gênero Atya Leach, Atys Leach, 1815:345. Atya Leach, 1816:421. (Leach, 1815) ] Espécie tipo.- Atys scaber Leach, 1815:345 [i gu al a Atya scabra Diagnose.- Rostro achatado lateralmente, com ou sem espinhos em sua margem dorsal e margem ventral com ou sem dentes. Carapaça com espinhos antenal e pterigostômico. Margem ventral da segunda à quinta pleura abdominal com ou sem dentículos. Pereópodes sem exópodes. Primeiro e segundo pares de pereópodes com carpo distintamente mais largo do que longo, sendo mais curto que os dedos; com uma escavação na sua extremidade distal, que permite o encaixe do própode; dedos das quelas terminando em longos pincéis de pêlos. Três últimos pares de pereópodes cobertos de tubérculos esclerosados; superfície flexora do dátilo com tubérculos agrupados ou arranjados em uma fileira longitudinal. Telso alargado e com sua extremidade distal arredondada. Observações.- Espécimens deste gênero habitam rios de correnteza, onde há muito oxigênio dissolvido, e com fundo de pedras. Os tubérculos presentes nos três últimos pares de pereópodes, sobretudo no terceiro que é mais forte, representam importante adaptação, permitindo que o animal se agarre ao fundo e não seja levado pela correnteza (HOBBS & HART, 1982).
29 19 Atya gabonensis Giebel, Est. I (a-h) Atya gabonensis Giebel, 1875: Thompson, 1901:22 - Bouvier, 1904: : 110, 112, 123, 124, :20, 22, 26, 27, 29, 293, Monod, 1928: : :110, 119, : 1203, :375 - Oliveira, 1945:179 - Holthuis, 1951:9, :69, 71, Hobbs, 1980:111 - Hobbs & Hart, 1982:48 - Holthuis, 1993:3. Euatya sculptilis Koelbel, 1884:317 - Hobbs, 1980: 111. Atya sculptata Ortmann, 1890:465. Atya (Atya) gabonensis - Ortmann, 1895:410, 414, : 184, 185, 186. Euatya sculptilis - Ortmann, 1897: 185. Atya scabra - Rathbun, 1900:313. Caracterização.- Rostro com carena mediana, sem dentes em suas margens dorsal e ventral, apresentando projeções laterais que formam, com o rostro, um ângulo nunca obtuso. Carapaça fortemente esculpida e com espinho pterigostômico pouco distinto. Pleuras abdominais sem dentículos esclerosados na sua margem ventral. Esterno do quinto segmento abdominal com projeção em forma de chifre. Telso com fileiras pares de 4 a 5 espinhos dorsais. Carena preanal com extremidade aguda e não direcionada para o telso. Apêndice masculino no segundo par de pleópodes do macho, 2.4 vezes mais longo do que apêndice interno. Primeiro par de pereópodes alcança o último artículo do pedúnculo antenular. Segundo par de pereópodes alcança os dedos do primeiro par de pereópodes. Primeiro e segundo pares de pereópodes apresentam o carpo mais largo do que longo e seus dedos terminam em longos pincéis de pêlos.
30 20 Terceiro par de pereópodes ultrapassa o último artículo do pedúnculo antenular com o carpo inteiro. Mero um pouco achatado ventralmente e 2.5 mais longo do que largo, superficie lateral com fileiras irregulares de tubérculos córneos com extremidades achatadas e em forma de escama; apresenta um tubérculo na superficie médio ventral do mero em oposição a um outro tubérculo do carpo. Mero 2.4 vezes o comprimento do carpo. Própode 2 vezes o comprimento do carpo, superficie extensora do própode com tubérculos córneos semelhantes aos do mero e com poucos largamente espalhados na sua superficie flexora. Superficie flexora do dátilo com um grupo de 4 a 5 tubérculos próximos à extremidade córnea. Diaresis do ramo externo de cada urópode, marcada por uma fileira de 17 a 19 dentículos articulados, e terminando em um espinho fixo um pouco maior. Tamanho.- O comprimento dos machos varia de 135 mm a 156 mm, e o da fêmea ovada é de 101 mm. Os ovos são numerosos e pequenos. Material examinado: DZUFRJ-3541, 1 macho - Natividade de Carangola, Fazenda São José (200 a 300 metros de altitude). DZUFRJ-3540, 3 fêmeas (1 ovada) - Natividade de Carangola, Fazenda São José (200 a 300 metros de altitude). MNRJ-2696, 1 macho - São Fidélis, Pureza, rio Paraíba do Sul. MNRJ-2694, 1 macho - Itaocara, rio Paraíba do Sul. MNRJ-2699, 1 macho - Itaocara (a montante de Coronel Teixeira), no Paraíba do Sul. MZUSP-10288, 1 macho - Itaocara, Coronel Teixeira, rio Paraíba do Sul. MZUSP-10289, 2 machos - Itaocara, Coronel Teixeira, rio Paraíba do Sul.
31 21 Localidade-tipo.- Gabão, África. O tipo foi depositado no British Museum (Natural Histmy ), Londres, Inglaterra. Distribuição geográfica.- África: do Senegal até o Zaire; América do Sul: Suriname, nordeste e sudeste do Brasil, México. Observações.- Os adultos desta espécie habitam rios de correnteza rápida e com fundo de pedras. Os indivíduos jovens podem ser capturados entre a vegetação (HOBBS & HART, 1982). Foram encontrados em altitudes de até 300 metros. Segundo o mesmo autor, a distribuição geográfica de A. gabonensis era limitada à costa oeste da África, Suriname e nordeste do Brasil (Estado do Piauí). Em 1994, sua distribuição foi ampliada para o norte do Estado do Rio de Janeiro, na bacia do Paraíba do Sul (FONSECA, OSTROVSKI & SILVA- FERREIRA, 1994). Atya scabra (Leach, 1815) Est. II (a-h) Atys scaber Leach, 1815:345. Atya mexicana Wiegmann, 1836:145. Ataya margaritacea Milne-Edwards, 1864: 148 (grafia errada). Atya punctata Kingsley, 1878:91.
32 22 Atya scabra - Rathbun, 1901: Bouvier, 1925:314 - Villalobos, 1943:7 - Holthuis, 1966:234 - Fausto-Filho, 1968:28 - Chace, 1972: 14 - Abele, 1975:56, 57 - Escobar, 1977:126 - Rodriguez, 1981:46 - Hobbs & Hart, 1982:106 - Coelho & Ramos-Porto, 1985:404 - Abele & Kim, 1989:4. Caracterização.- Rostro curto sem dentes em suas margens dorsal e ventral e com projeções laterais formando ângulo obtuso com o rostro. Superficie dorsal com carena mediana um tanto afiada. Carapaça fracamente esculpida, podendo ser nodosa em indivíduos maiores. Espinhos lateral e pterigostômico fortes. Margem ventral da segunda até a quinta pleura com uma fileira de dentículos esclerosados. Esternito do quinto segmento abdominal com projeção mediana comprimida e nunca formando estrutura em forma de chifre, ultrapassando o sexto esternito abdominal. Carena preanal com extremidade aguda ou arredondada e dirigida para trás em direção ao telso. Telso com 2 fileiras de 5 a 7 espinhos dorsais. Apêndice masculino no segundo par de pleópodes do macho, 3.4 vezes mais longo do que o apêndice interno. Primeiro e segundo pereópodes com dedos terminando em longos pincéis de pêlos. Primeiro pereópode atinge além da extremidade do último artículo antenular com extremidade do pincel de pêlos. Segundo pereópode com carpo mais largo do que longo. Terceiro pereópode ultrapassando o último artículo do pedúnculo antenular com o carpo inteiro. Mero arredondado ventralmente e 2.6 vezes mais longo do que largo; superficie lateral com fileiras irregulares de tubérculos córneos. Carpo tem a metade do comprimento do mero. Própode 1.4 vezes mais longo do que o carpo; com fileiras de tubérculos esclerosados, estando os da superficie flexora arranjados em duas fileiras longitudinais. Dátilo com uma única fileira longitudinal de 6 a 7 tubérculos na sua superficie flexora.
33 23 Diaresis do ramo externo de cada urópode, marcada por uma fileira de 15 a 20 dentículos articulados e tenninando em um espinho fixo. Tamanho.- O comprimento dos machos examinados varia de 55 a 59 mm e o das fêmeas examinados fica entre 24 e 36 mm. A fêmea ovada tem comprimento de 40 mm. Os ovos são numerosos e pequenos. Material examinado: DZUFRJ-2041, 1 macho - Natividade de Carangola, Fazenda São José. MNRJ-2698, 1 macho - Campo Grande, rio da Prata. DZUFRJ-2113, 2 fêmeas - Mangaratiba, rio Grande. MNRJ-2695, 1 macho - Angra dos Reis, rio Zungu. DZUFRJ-2106, 1 fêmea - Parati, rio São Roque. DZUFRJ-2097, 1 macho - Parati, rio Parati-Mirim. MNRJ-4162, 1 fêmea - Parati, afluente da margem esquerda do rio Parati Mirim. Localidade tipo.- Foi estabelecido neótipo para Misantla, Estado de Vera Cruz, México. O neótipo foi depositado no British Museum (Natural History), Londres, Inglaterra. Distribuição geográfica. - (México, Antilhas e Brasil até Santa Catarina). África Ocidental, Austrália e Américas
34 24 Observações.- Os indivíduos desta espécie habitam águas doces e rasas, com correnteza e fundo de pedras, escondendo-se debaixo de pedras e folhas. Tem uma ampla distribuição no rio. Os indivíduos desta espécie passam as primeiras fases de seu desenvolvimento perto da costa, posteriormente mi gr ando rio acima para distribuir-se em áreas muito distantes do estuário (VILLALOBOS, 1959). Foram encontrados em altitudes de O a 300 metros. Gênero Potimirim Holthuis, Villalobos, Car. Emend. Potimirim Holthuis, 1954:2 - Chace & Hobbs, 1969:76 - Chace, 1972: 14 - Rodriguez, 1980: :41 - Villalobos, 1982:223 - Abele & Kim, 1989:5. Espécie tipo: Caridina mexicana De Saussure, Diagnose.- Rostro curto, sem dentes na margem dorsal e com dentes na margem ventral. Espinhos supra orbitais ausentes; espinho antenal presente e espinho pterigostômico distintamente presente nas fêmeas, podendo ser fraco ou ausente nos machos. Pereópodes sem exópodes em sua base. Epipoditos presentes nos três ou quatro primeiros pares de pereópodes. Dois primeiros pares de pereópodes com os dedos de suas quelas terminando em pincéis de longos pêlos. Carpos do primeiro e segundo pares de pereópodes com extremidade distal escavada onde se encaixa a extremidade proximal do própode; segundo par de pereópodes
35 com carpo mais longo do que largo. Segundo par de pleópodes do macho com apêndice masculino, estrutura importante como característica taxonômica. 25 Potimirim glabra (Kingsley, 1878) Est. III (a-d) Atyoida glabra Kingsley, 1878:93. Potimirim glabra; Holthuis, 1954:3, fig a: Villalobos, 1960:273 - Smalley, 1963: 177, fig. 1 - Chace & Hobbs, 1969:76, figs. 15, 19 b, c - Abele & Blum, 1977:244 - Hart, 1980:845 - Rodriguez, 1981:43,46 - Villalobos, 1982:217 - Felgenhauer & Abele, 1985:401 - Holthuis, 1986:445 -Abele & Kim, Potimirim brasiliana Villalobos, 1959:275, lams. I-IV. Caracterização.- Rostro ultrapassando a extremidade distal do primeiro artículo do pedúnculo antenular, margem dorsal do rostro sem dentes e com sua extremidade um pouco voltada para baixo; margem ventral com 2 ou 3 dentes, excepcionalmente 1. Ângulo pterigostômico dos machos pode ser arredondado ou agudo, mas nunca formando espinho; e nas fêmeas sempre agudo. Pleuras abdominais arredondadas. Pleura do quinto somito abdominal das fêmeas com a extremidade terminando em um vértice arredondado. Telso com fileiras pares de 4 a 6 espinhos dorsais. Epipoditos nos 4 primeiros pares de pereópodes. Primeiro par de pereópodes com carpo mais curto que os dedos. Segundo par de pereópodes com carpo distintamente mais longo do que largo, e
36 26 quase tão longo quanto os dedos. Dátilo do terceiro par de pereópodes apresentando 7 a 8 dentes (incluindo o terminal) em seu bordo inferior, os quais são encurvados, com sua extremidade voltada pará trás; carpo com pequenos e numerosos tubérculos na região inf eroproximal e também com 2 espinhos, um em cada lado da mesma região, os quais correspondem aos órgãos sexuais caracteristicos dos machos; mero 3.5 vezes mais longo do que largo, com metade distal alargada, provida de urna série longitudinal de 4 espinhos e também com numerosos e pequenos tubérculos em sua margem inferior. Pereópodes do quarto par com dátilo apresentando a margem inferior com 7 dentes terminando em um mais forte. Pereópodes do quinto par têm dátilo com margem inferior pectinada terminando em 2 dentes fortes. Apêndice masculino do segundo par de pleópodes com margem posterior dividida em 3 lobos, estando o proximal separado dos outros 2 por urna depressão sem espinhos. Tamanho.- O comprimento dos machos varia de 11 a 26 mm e o das fêmeas fica entre 11 e 19 mm. A maior fêmea ovada tem 27 mm de comprimento. Ovos numerosos e pequenos. Material examinado: MNRJ-4401, 16 fêmeas (ovadas) - Itabapoana, Barra de Itabapoana (próximo à ponte RJ/ES). MNRJ-2684, 3 fêmeas (2 ovadas) - Araruama, lagoa de Juturnaíba. DZUFRJ-2133, 3 machos - Miguel Pereira, cachoeira do rio Santa Branca (BR 125) MNRJ-2715, 10 machos e 13 fêmeas - Ilha de Itacuruçá (riacho do lado esquerdo da praia Grande) DZUFRJ-2120, 296 machos e 98 fêmeas - Mangaratiba, rio Grande.
37 27 DZUFRJ-2122, 2 fêmeas (ovadas) - Mangaratiba, serra do Piloto (aprox. 500 m de altitude) MNRJ-2719, 3 machos e l fêmea (ovada) - Ponta da Figueira (entre Mangaratiba e Conceição do J acareí). DZUFRJ-2125, 3 fêmeas (ovadas) - Lídice, afluente do rio da Guarda. MNRJ-2686, 4 machos e 3 fêmeas (ovadas) - Angra dos Reis, rio Ariró Pequeno. MNRJ-2745, 3 fêmeas -Angra dos Reis, Ilha Grande. DZUFRJ-2067, 15 machos e 6 fêmeas - Angra dos Reis, Ilha Grande. MNRJ-2734, 53 machos e 64 fêmeas (14 ovadas) - Parati, riacho da prainha entre Mambucaba e T arituba. DZUFRJ-2099, 11 machos - Parati, rio São Roque. DZUFRJ-2101, 1 macho e 1 fêmea (1 ovada) - Parati, rio São Roque. DZUFRJ-2077, 3 machos e 6 fêmeas (2 ovadas) - Parati, rio São Roque. DZUFRJ-2080, 2 fêmeas (1 ovada) - Parati, rio São Roque. DZUFRJ-2074, 2 fêmeas (ovadas) - Parati, rio São Roque. DZUFRJ-2081, 9 fêmeas (ovadas) - Parati, rio Parati-Mirim. DZUFRJ-2083, 2 fêmeas - Parati, rio Parati-Mirim. DZUFRJ-2085, l macho e l fêmea (ovada) - Parati, rio Parati-Mirim. DZUFRJ-2082, 2 fêmeas (1 ovada) - Parati, rio Graúna. MZUSP-7373, l macho - Parati. Localidade tipo.- Costa oeste da República da Nicarágua. O tipo foi depositado no Museum of Comparative Zoology, Harvard University, Cambridge, Massachusets, EUA. Distribuição geográfica.- Oeste do México até o Equador e do leste do México até São Paulo (Brasil).
38 28 Observações.- São habitantes de águas doces, em rios com fundo de cascalho e com correnteza moderada a rápida (CHACE & HOBBS, 1969). Foram encontrados em altitudes de até 500 metros (Serra do Piloto, Mangaratiba), estando junto à vegetação marginal e também associados às raízes de macrófitas. As fêmeas de P. glabra podem, às vezes, ser confundidas com as de P. potimirim (MÜLLER 1881), das quais podem ser diferenciadas pela presença de epipodito no quarto par de pereópodes e também pela largura do mero do terceiro pereópode, que é maior em P. glabra. Potimirim potimirim (Müller, 1881) Est. IV ( a-d). Atyoida potimirim Müller, 1881: 117. Ortmania potimirim; Rathbun, 1901: 120. Ortmania mexicana; Bouvier, 1925:284. Potimirim mexicana; Holthuis, 1954:4. Potimirim potimirim; Villalobos, 1959: , lams. X-XII - Abele, 1972:215, fig. 4 - Rodriguez, 1981:46 - Villalobos, 1982:217 -Abele & Kim, 1986: Caracterização.- Rostro alcançando a porção proximal do segundo artículo do pedúnculo antenular, margem dorsal do rostro quase reta e sem dentes;
39 29 margem ventral com 2 ou 3 dentes. Ângulo pterigostômico dos machos sem espinho; fêmeas com um espinho pterigostômico agudo. Pleura do quinto somito abdominal das fêmeas com extremidade terminando em ângulo reto. Telso com fileiras pares de 5 espinhos dorsais. Epipodito nos 3 primeiros pares de pereópodes. Primeiro par de pereópodes com carpo mais curto que os dedos. Segundo par de pereópodes com carpo distintamente mais longo do que largo, e quase tão longo quanto os dedos. Pereópodes do terceiro par têm dátilo com 7 a 8 dentes_ (incluindo o terminal) na margem inferior; carpo com 2 espinhos, um em cada lado da região inf eroproximal, os quais correspondem aos órgãos sexuais característicos dos machos; mero um pouco alargado na metade distal, sendo 4.75 vezes tão longo quanto largo, e com uma fileira longitudinal de 4 espinhos na margem inferior. Pereópodes do quarto par têm dátilo com 7 a 8 dentes (incluindo o terminal) na sua margem inferior, com extremidade um pouco voltada para trás. Carpo com 2 espinhos lateroproximais e um espinho mais distal na superfície externa; possui também pequenos e numerosos tubérculos na superfície inferior da sua porção proximal; mero com uma série de 4 espinhos na sua metade distal. Pereópodes do quinto par têm dátilo com margem inferior pectinada e terminando em 2 espinhos fortes. Apêndice masculino do segundo par de pleópodes com margem posterior dividida em 3 lobos estando o proximal separado dos outros 2 por uma depressão provida de espinhos, e menos acentuada que em P. glabra. Tamanho.- O comprimento dos machos examinados varia de 11 a 25 mm e o das fêmeas examinadas fica entre 11 a 24 mm. A maior fêmea ovada examinada tem 21 mm de comprimento. Ovos numerosos e pequenos.
40 30 Material examinado: DZUFRJ-2054, 3 fêmeas - São Fidélis, rio Dois Rios (afluente do rio Paraíba do Sul). DZUFRJ-2055, 2 fêmeas - São Fidélis, Cambiasca. DZUFRJ-2040, 3 fêmeas (ovadas) - Maricá, rio Bananal. MNRJ-2664, 2 fêmeas (ovadas) - rio em Muriqui. MNRJ-27q, 12 machos e 12 fêmeas (2 ovadas) - Angra dos Reis, no Florestão. DZUFRJ-2066, 10 machos e 17 fêmeas - Angra dos Reis, rio Bracuí. MNRJ-898, 1 fêmea - Angra dos Reis, rio Bracuí. MNRJ-2736, 4 machos e 12 fêmeas (6 ovadas) - Angra dos Reis, rio Grataú. DZUFRJ-2071, 2 machos - Parati, rio São Roque. MZUSP-11295, 2 fêmeas (1 ovada) - Parati, rio Barra Grande. DZUFRJ-2075, 3 fêmeas (ovadas) - Parati, rio Parati-Mirim. DZUFRJ-2076, 10 machos e 3 fêmeas (2 ovadas) - Parati, rio Parati-Mirim. MNRJ-27 16, 15 machos e 20 fêmeas - Parati, rio Parati-Mirim. MNRJ-4164, 6 machos e 13 fêmeas (ovadas) - Parati, rio Parati-Mirim (local com influencia de maré) MNRJ-4163, 1 fêmea - Parati, afluente da margem direita do rio Parati Mirim. Localidade tipo.- Rio Itajaí, Santa Catarina, Brasil. Distribuição geográfica.- Flórida (EUA) até Santa Catarina (Brasil). Observações.- Apesar de serem habitantes de águas doces, é possível encontrar espécimens de P. potimirim no mar, a uma certa distância da costa. Alguns indivíduos encontram-se associados às raízes de macrófitas que são
41 arrastadas pelos rios, permitindo uma possível dispersão desta espécie, assim como de outras do gênero Potimirim (VILLALOBOS, 1959). 31 P. potimirim é muito semelhante à P. mexicana (De Saussure, 1857), que ocorre no México, Costa Rica, Jamaica e Porto Rico. P. potimirim tem o apêndice masculino mais estreito e as depressões entre os lobos, na margem posterior, são mais profundas. Segundo COELHO & RAMOS-PORTO (1985) são animais encontrados em altitudes de até 100 metros. Família Palaemonidae Rafinesque, 1815 Palaemonidae Rafinesque, Holthuis, 1950: : :42 - Chace & Hobbs, 1969:87 - Chace, 1972:17 - Gomes-Corrêa, 1977:16 - Rodriguez, 1980:85 - Williams, 1984:63. Diagnose.- Rostro geralmente bem desenvolvido, lateralmente comprimido, com dentes na margem dorsal e com ou sem dentes na margem ventral. Carapaça com espinho antenal sempre presente e com ou sem espinho hepático ou branquiostegal. Sulco branquiostegal presente ou ausente. Exopoditos presentes em todos os maxilípedes. Terceiros maxilípedes filiformes, com endopodito pluriarticulado e exopodito inteiro. Pereópodes sem exopoditos. Dois primeiros pares de pereópodes quelados e com carpo inteiro, pereópodes do segundo par geralmente maiores do que os do primeiro. Três últimos pares de pereópodes com
42 dátilo simples ou bífido. Apêndice masculino do segundo par de pleópodes podendo estar presente ou ausente. 32 A família compreende quatro subfamílias, das quais apenas a subfamília Palaemoninae possui representantes na rede fluvial do Estado do Rio de Janeiro. Subfamília Palaemoninae Dana, 1852 Palaemoninae Dana, 1852: 17, 24 - Holthuis, 1950:4-1952:1-1955:43 - Williams, 1965:50 - Chace & Hobbs, 1969:87 - Chace, 1972: 18 - Gomes-Corrêa, 1977:33 - Williams, 1984:63. Diagnose.- Rostro com dentes nas margens dorsal e ventral. Carapaça com espinho antenal sempre presente e com ou sem espinho hepático ou branquiostegal; sulco branquiostegal presente ou ausente. Telso com margem posterior provida de 2 pares de espinhos e um ou mais pares de cerdas. Flagelo antenular superior com os ramos fusionados na porção basal. Mandíbulas com ou sem palpo. Segundo par de pereópodes mais forte do que o primeiro. Apêndice masculino sempre presente no segundo pleópode.
43 CHAVE PARA AS ESPÉCIES DA FAMÍLIA PALAEMONIDAE DO ESTADO DO RIO DE JANEIRO Espinho hepático ausente. Espinho branquiostegal presente Pa/aemon (Palaemon) panda/iformis ( est. V) Espinho hepático presente. Espinho branquiostegal ausente Rostro longo, ultrapassando a extremidade distal do escafocerito. Dedos das quelas do 2 º par de pereópodes cobertos de pêlos Macrobrachium acanthurus ( est. VI) Rostro curto, alcançando a extremidade distal do pedúnculo antenular ou mais curto. Dedos das quelas do 2 par de pereópodes sem pêlos e com fortes espinhos Margem posterior do telso com uma extremidade truncada e com espinhos posteriores curtos. Pereópodes do segundo par com o carpo menor que o mero... Macrobrachium carcinus (est. VII) Margem posterior do telso com uma distinta extremidade aguda e com espinhos posteriores internos tão ou mais longos que a citada extremidade. Pereópodes do segundo par com o carpo tão ou mais longo quanto o mero Pereópodes do segundo par muito desiguais em forma e tamanho e com própode inchado. Margem dorsal do rostro com 12 a 15 dentes Macrobrachium olfersii (est. VIII)
44 34 Pereópodes do segundo par iguais em forma, podendo ser desiguais ou ligeiramente desiguais em tamanho e com própode nunca inchado. Margem dorsal do rostro com 5 a 10 dentes Segundo par de pereópodes com dedos deixando um espaço quando fechados. Espinhos internos do telso ultrapassando a extremidade do mesmo Macrobrachium potiuna ( est. IX) Segundo pereópode com dedos não deixando espaço quando fechados. Espinhos internos do telso ultrapassando a extremidade do mesmo Macrobrachium iheringi (est. X) Gênero Palaemon Weber, Palaemon Weber, 1795:94 - Fabricius, 1798:378, Holthuis, 1952: :46. Espécie tipo.- Cancer Squila Linnaeus, Diagnose. - Rostro longo e com dentes em suas margens dorsal e ventral. Carapaça com espinhos antenal e branquiostegal, sem espinho hepático e com sulco branquiostegal. Mandíbula com palpo bi ou triarticulado. Dois primeiros pares de pereópodes delgados, sendo o segundo ligeiramente mais forte que o primeiro. Três últimos pares de pereópodes com dátilo simples. Telso terminando em uma extremidade aguda.
45 35 Palaemon (Palaemon) pandaliformis (Stimpson, 1871). Est. V (a-d). Leander pandaliformis Stimpson, 1871:130. Leander potitinga Ortmann, 1897: 193, pl. 1, fig Thering, 1897:422 - Moreira, 1901: 12. Palaemon (Palaemon) pandaliformis; Holthuis, 1950:7-1952: 187, pl. 46, figs. g. 1 - Chace & Hobbs, 1969: 111, figs. 26, 28 a - Chace, 1972:21 - Gomes-Corrêa, 1977:103, est. XXVI, figs Hart, 1980:845 - Rodriguez, 1980:128, :43, 47 - Villalobos, 1982:218, 225. Caracterização.- Rostro longo e delgado, ultrapassando a extremidade distal do escaf ocerito, margem dorsal quase reta ou com a extremidade voltada para cima; margem dorsal com 1 ou 2 dentes subapicais, um espaço desarmado e 5 a 7 dentes na metade proximal, sendo que um está localizado sobre a carapaça, atrás do nível da órbita; margem ventral com 5 a 8 dentes. Carapaça sem espinho hepático e com espinho branquiostegal presente em sua margem anterior. Carapaça e abdome lisos. Pleura do quinto somito abdominal com extremidade terminando em vértice arredondado. Sexto somito abdominal quase o dobro do quinto e tão longo quanto o telso. Telso um tanto delgado e com 2 pares de espinhos dorsais; margem posterior com extremidade aguda, e com 2 pares de espinhos posteriores, sendo que os internos ultrapassam a extremidade do telso. Mandíbula apresentando palpo triarticulado.
46 36 Primeiro par de pereópodes alcançando a extremidade distal do escafocerito, dedos um pouco mais longos que a palma. Segundo par de pereópodes iguais na forma e no tamanho e bastante delgados; ultrapassa o escafocerito com metade da palma quando esticado; dedos um pouco mais curtos que a palma; margem cortante do dedo fixo lisa e a do dátilo com um pequeno dente na porção proximal; carpo 2.5 vezes o comprimento da palma e 1.5 vez o comprimento do mero. Terceiro par de pereópodes alcançando a extremidade distal do escafocerito; própode 2.5 vezes mais longo do que o dátilo, 1.5 vez mais longo do que o carpo e um pouco menor que o mero. Quarto par de pereópodes ultrapassa a extremidade distal do escafocerito com o dátilo inteiro; própode 2.5 vezes tão longo quanto o dátilo, quase 2 vezes mais longo do que o dátilo e um pouco mais longo que o mero. Quinto par de pereópodes ultrapassa o escafocerito com metade do própode; própode 3.5 vezes mais longo do que o dátilo, 2 vezes mais longo do que o carpo e um pouco maior que o mero. Tamanho.- O comprimento dos machos examinados varia de 7 a 36 mm. A maior fêmea ovada examinada tem 40 mm de comprimento total. Os ovos são numerosos e pequenos. Material examinado: DZUFRJ-2056, 18 machos e 23 fêmeas - Campos, canal da lagoa da Cataia. DZUFRJ-2058, 3 machos - Campos, canal da lagoa da Cataia. DZUFRJ-2057, 2 machos - Campos, canal da lagoa da Cataia. MZUSP-1955, 7 machos - rio Paraíba do Sul. MZUSP-1956, 7 machos e 12 fêmeas (10 ovadas) - rio Paraíba do Sul. LAB. LIMNOLOGIA-UFRJ, 1 fêmea (ovada) - Macaé, lagoa Comprida.
47 37 MNRJ-1046, 2 machos e 7 fêmeas (2 ovadas) - Duque de Caxias, rio Paô. Col. em 27/06/1941. MNRJ-1040, 1 macho e 9 fêmeas (7 ovadas) - Rio de Janeiro, rio Pavuna. Col. em 20/07/1944. MNRJ-998, 3 fêmeas (ovadas) - Rio de Janeiro, Barra da Tijuca, canal do Cortado MNRJ-1039, 8 machos e 62 fêmeas (1 ovada) - Rio de Janeiro, Recreio dos Bandeirantes, lagoa de Marapendi. Col. em 05/10/1961. MNRJ-1042, 4 machos e 14 fêmeas (13 ovadas) - Itacuruçá, rio Itin gu açu. DZUFRJ-3788, 4 machos - Mangaratiba, Fazenda Lindo Vale, praia da Cruz. DZUFRJ-2068, 1 macho - Angra dos Reis, rio Areia do Pontal. MNRJ-893, 9 machos e 2 fêmeas - Angra dos Reis, rio Saco. MNRJ-976, 4 machos e 15 fêmeas (1 ovada) -Angra dos Reis, rio do Frade. MNRJ-4402, 1 macho e 2 fêmeas - rio Mambucaba. DZUFRJ-2084, 2 machos e 5 fêmeas - Parati, rio Barra Grande. Localidade tipo.- Trinidad (Venezuela). O tipo foi depositado na coleção do United States National Museum, Washington, D.C., EUA. (Brasil). Distribuição geográfica.- Antilhas e Guatemala até Santa Catarina Observações.- São predominantemente de á gu a salobra, sendo encontrados próximo a foz dos rios. De acordo com COELHO & RAMOS-PORTO ( 1985) são encontrados em altitudes de O a 100 metros. Palaemon (Palaemon) panda/iformis mostra-se bastante relacionada à espécie Pa/aemon (Palaemon) graci/is (SMITH, 1871) da costa ocidental da
48 América, diferenciando-se dela pela forma da extremidade do telso e comprimento dos seus espinhos posteriores internos. 38 Gênero Macrobrachium Bate, Macrobrachium Bate, 1868:363 - Holthuis, 1950: : :53 - Chace & Hobbs, 1969:89 - Rodriguez, 1980: 113. Espécie tipo.- Macrobrachium americanum Bate, Diagnose.- Rostro bem desenvolvido, comprimido lateralmente e com dentes nas margens superior e inferior. Flagelo antenular superior com dois ramos fusionados na base. Carapaça com espinhos antenal e hepático presentes e espinho branquiostegal ausente. Sulco branquiostegal bem marcado. Maxilípedes com exopoditos. Mandíbula com palpo triarticulado. Primeiro par de pleópodes do macho sem apêndice interno. Telso com dois pares de espinhos dorsais e dois pares de espinhos posteriores. Macrobrachium acanthurus (Wiegmann, 1836). Est. VI (a-e). Palaemon acanthurus Wiegmann, 1836: 150.
49 39 Macrobrachium acanthurus; Pearse, 1911: Holthuis, 1952:45, pl. 8-9, figs.a, b :91 - Williams, 1965:52, fig Chace, 1972:20 - Gomes-Corrêa, 1977:60, est. XIII, figs Williams, 1984:66, figs Abele & Kim, 1989:5. Caracterização.- Rostro longo, geralmente alcançando ou ultrapassando a extremidade distal do escafocerito; margem dorsal com 9 a 11 dentes, 2 primeiros dentes sobre a carapaça, atrás da margem orbital, sendo o primeiro separado do segundo por um espaço maior do que entre o segundo e o terceiro; margem ventral com 4 a 7 dentes. Tanto na margem dorsal quanto na ventral os dentes proximais mostram-se mais juntos que os distais. Carapaça lisa, exceto por alguns eventuais pelos curtos na região anterolateral. Abdome liso. Pleura do quinto somito abdominal com extremidade terminando em vértice a gu do. Telso com margem posterior terminando em uma extremidade a gu da, flanqueada por dois pares de espinhos, sendo os mais internos mais longos que a referida extremidade. Primeiro par de pereópodes alcança a extremidade do escafocerito com o própode inteiro e, às vezes com parte do carpo; dedos mais curtos que a palma e carpo 2.5 vezes mais longo do que o própode. Segundo par de pereópodes delgados e i gu ais em forma e tamanho; alcançam a extremidade do escafocerito com o carpo inteiro ou uma parte do mero. Dedos não deixando espaço quando fechados, quase tão longos quanto a palma e cobertos por uma densa pubescência aveludada; margens cortantes armadas com um forte dente proximal no dátilo, outro no dedo fixo ( o dente do dátilo está localizado um pouco antes do dente do dedo fixo) e 3 ou 4 dentículos basais; própode 2 vezes mais longo do que o dátilo; carpo mais longo que a palma e que o mero. Terceiro par de pereópodes ultrapassa o escafocerito
50 40 com o própode inteiro e uma pequena parte do dátilo; própode 2.6 vezes mais longo do que o dátilo, 1.6 a 2 vezes do que o carpo e um pouco menor que o mero. Quarto par pereópodes alcança o escafocerito com a extremidade do dátilo; própode quase 3 vezes mais longo do que o dátilo, 1.5 do que o carpo e quase tão longo quanto o mero. Quinto par de pereópodes ultrapassa o escafocerito com uma pequena parte do dátilo; própode 3 vezes mais longo do que o dátilo, 1.6 a 2 vezes do que o carpo e tão longo quanto o mero ou um pouco mais curto. Tamanho.- O comprimento dos machos varia de 33 a 180 mm e o das fêmeas fica entre 26 a 96 mm. A maior fêmea ovada tem comprimento total de 11 O mm. Os ovos são numerosos e pequenos. Material examinado: MNRJ-3367, 1 macho - Barra de Itabapoana. MNRJ-4399, 2 fêmeas - Barra da Itabapoana, próximo à ponte RJ/ES. MZUSP-1929, 23 machos e 3 fêmeas (1 ovada) - rio Paraíba do Sul. MZUSP-1921, 22 machos e 2 fêmeas (1 ovada) - rio Paraíba do Sul. MZUSP-1926, 2 machos e 2 fêmeas (ovadas) - rio Paraíba do Sul. MZUSP-1934, 4 machos e 8 fêmeas (5 ovadas) - rio Paraíba do Sul. MZUSP-1935, 13 machos e 1 fêmea (ovada) - rio Paraíba do Sul. MZUSP-628, 10 machos e 9 fêmeas (3 ovadas) - São João da Barra. MNRJ-964, 1 macho e 1 fêmea - Campos, Atafona. MZUSP-1911, 1 macho - Campos, Atafona. MZUSP-6473, 1 macho e 4 fêmeas - Campos, Atafona. MNRJ-875, 1 macho - Campos, rio Novo, lagoa Feia. MNRJ-882, 3 machos e 3 fêmeas - Macaé, lagoa adjacente à Quissamã.
51 41 MNRJ-4160, 2 machos e 1 fêmea - Rio das Ostras, riacho que desemboca na praia do Mar do Norte. DZUFRJ-2033, 1 macho e 1 fêmea - Bom Jardim, rio São José (afluente do rio Grande). MNRJ-880, 3 machos - Araruama, lagoa de Jutumaíba. DZUFRJ-2026, 1 macho - Maricá, rio Engenho Novo. DZUFRJ-2028, 1 macho - Maricá, lagoa de Maricá. MZUSP-1932, 1 macho - Maricá, lagoa de Maricá. DZUFRJ-3568, 1 macho - Maricá, laguna de Maricá. MNRJ-914, 3 machos e 2 fêmeas - Niterói, Jurujuba. MNRJ-932, 2 machos e 3 fêmeas - Niterói, Jurujuba. MNRJ-916, 2 machos - Duque de Caxias. Col. Dez/1941. MNRJ-883, 2 machos e 2 fêmeas - Rio de Janeiro, lagoa Rodrigo de Freitas. MNRJ-958, 1 fêmea - Rio de Janeiro, lagoa de Jacarepaguá. MNRJ-927, 1 macho - Rio de Janeiro, Jacarepaguá, rio Camorim. Col. 23/06/1968. MNRJ-961, 1 fêmea - Pedra de Guaratiba. MNRJ-963, 1 macho e l fêmea - rio em Guaratiba. MNRJ-4398, 2 fêmeas - Barra de Guaratiba, riacho na estrada da Ilha. MNRJ-922, 1 fêmea (ovada) - Rio de Janeiro, rio Piraquê. MNRJ-872, 1 fêmea (ovada) - Sepetiba, praia de D. Luísa. MNRJ-920, 10 machos e 7 fêmeas - Itacuruçá, rio Itinguaçu. MNRJ-923, 5 machos e 9 fêmeas (3 ovadas) - Ilha da Marambaia. MNRJ-918, 1 macho - Ilha da Marambaia. MNRJ-921, 2 machos e 4 fêmeas (2 ovadas) - Mangaratiba, praia do Saco. DZUFRJ-2123, 1 macho - Mangaratiba, Fazenda Lindo Vale, praia da Cruz. DZUFRJ-2117, 9 machos e 15 fêmeas (11 ovadas) - Mangaratiba, Fazenda Lindo Vale.
52 42 DZUFRJ-2112, 6 machos - Mangaratiba, Fazenda Lindo Vale. DZUFRJ-3784, 13 machos e 1 fêmea - Mangaratiba, Fazenda Lindo Vale. DZUFRJ-3789, 13 machos e 6 fêmeas (4 ovadas) - Mangaratiba, Fazenda Lindo Vale. DZUFRJ-3787, 6 machos e 21 fêmeas (1 ovada) - Mangaratiba, Fazenda Lindo Vale. DZUFRJ-3791, 12 machos e 1 fêmea - Mangaratiba, Fazenda Lindo Vale. DZUFRJ-3786, 4 machos e 1 fêmea - Mangaratiba, Fazenda Lindo Vale. DZUFRJ-2115, 1 macho - Mangaratiba, rio na praia do Saí. MNRJ-915, 4 machos e 5 fêmeas - Conceição do Jacareí, Tanque. MNRJ-3342, 2 machos e 1 fêmea (ovada) - Angra dos Reis, Ilha Grande, riacho na praia de Provetá. MZUSP-206, 1 macho -Angra dos Reis, Ilha Grande. MNRJ-917, 1 fêmea -Angra dos Reis, Japuíba. DZUFRJ-2059, 2 machos e 1 fêmea (ovada) - Angra dos Reis, Caputera, rio Areia do Pontal. MNRJ-977, 1 macho -Angra dos Reis, rio do Frade. MNRJ-919, 2 machos - Parati, Fazenda Laranjeiras. DZUFRJ-2110, 1 macho e 6 fêmeas (1 ovada) - Parati, rio Graúna. DZUFRJ-2092, 2 machos - Parati, rio São Roque. DZUFRJ-2073, 1 fêmea - Parati, rio São Roque. DZUFRJ-2073, 1 fêmea - Parati, rio São Roque. DZUFRJ-2096, 1 macho - Parati, rio Barra Grande. DZUFRJ-2103, 1 macho e 2 fêmeas - Parati, rio Barra Grande. DZUFRJ-2091, 1 fêmea (ovada) - Parati, rio Barra Grande. MNRJ-4166, 2 machos - Parati, rio Parati-Mirim (local com influência da maré). DZUFRJ-2109, 2 machos e 1 fêmea - Parati, rio Parati-Mirim.
53 43 DZUFRJ-2102, 4 machos - Parati, rio Parati-Mirim. DZUFRJ-2089, 1 macho e 5 fêmeas - Parati, rio Parati-Mirim. DZUFRJ-2087, 2 machos e 2 fêmeas - Parati, rio Parati-Mirim. Localidade tipo.- Costa brasileira. O tipo foi depositado no Zoologisch Museum, Berlim, Alemanha. Grande do Sul (Brasil). Distribuição geográfica.- Carolina do Norte (EUA), Antilhas até o Rio Observações.- Indivíduos desta espécie são habitantes de águas doces e salobras. De acordo com ESCOBAR ( 1979), podem ser encontrados preferencialmente em trechos de rio de correnteza suave e com fundo lodoso - arenoso, onde existe muito material vegetal em decomposição. Também podem ser capturados em remansos com fundo de pedra, e perto do estuário (fêmeas ovadas). O rostro pode variar em comprimento e formato. Fêmeas adultas e machos jovens têm o segundo par de pereópodes mais curto, mais delgado, menos espinuloso e pubescente que machos adultos. De acordo com ESCOBAR, 1979 são encontrados em altitudes de até 250 metros. Macrobrachium carcinus (Linnaeus, 1758). Est. VII (a-d). Potipema Marcgraf, 1648: 187.
54 44 Astacus jluviatilis major chelis acu/eatis Sloane, 1725 :271, pi. 245, fig. 2. Cancer carcinus Linnaeus, 1758:631. Macrobrachium jamaicense Pearse, 1915 :551. Macrobrachium carcinus - Hedgpeth, 1949:31, figs. lb, 3, 5 - Holthuis, 1952: 114, pi. 30, 31, figs. a, c :96, fig Chace & Hobbs, 1969:93, figs. 21, 25 b-h - Chace, 1972:20 - Coelho & Ramos-Porto, Gomes-Corrêa, 1977:43, est. VIII, figs Rodriguez, 1980:123, fig. 35 a-b - Williams, 1984:68 - Abele & Kim, 1989:6. Caracterização.- Rostro curto, alcançando a extremidade distal do pedúnculo antenular; margem dorsal sinuosa e extremidade um pouco voltada para cima; margem dorsal armada com 11 a 14 dentes distribuídos regularmente, dos quais 4 a 6 estão localizados sobre a carapaça, atrás da margem orbital. Carapaça lisa ou áspera na parte lateral, nos machos adultos. Abdome liso. Pleura do quinto somito abdominal com ângulo posterior reto e extremidade arredondada. Telso com espinhos posteriores curtos, os quais não ultrapassam sua extremidade truncada. Primeiro par de pereópodes alcança a extremidade distal do escafocerito com a quela inteira e, às vezes, com a metade do carpo. Segundo par de pereópodes, do macho adulto, fortes e iguais em forma e tamanho, com dedos um pouco mais curtos que a palma; dedos, quando cruzados, deixam um espaço entre eles na parte proximal; margens cortantes com grandes dentes, um na metade do comprimento do dátilo e um outro no final do terço proximal do dedo fixo; palma levemente comprimida, entre 3 a 4 vezes mais longa do que larga; carpo com mais ou menos a metade do comprimento da palma e um pouco menor que o mero. Terceiro par de pereópodes ultrapassa o escafocerito com o própode e uma parte do
55 45 dátilo; própode 2.5 vezes mais longo do que o dátilo, 1.3 a 1. 7 vezes do que o carpo e menor que o mero. Quarto par de pereópodes com própode 2.5 vezes mais longo do que o dátilo, 2 vezes mais do que o carpo e menor que o mero. Quinto par de pereópodes 2.5 vezes mais longo do que o dátilo, 1.4 vezes mais do que o carpo e menor que o mero. Tamanho.- O comprimento dos machos varia de 25 a 240 mm e o das fêmeas ovadas fica entre 24 e 170 mm. Os ovos são pequenos e numerosos. Material examinado: MNRJ-2672, 2 machos e 3 fêmeas (2 ovadas) - Barra de Itabapoana, rio Itabapoana. MZUSP-1918, 1 macho - rio Muriaé. MNRJ-2669, 6 machos e 7 fêmeas (3 ovadas) - São Fidélis, rio Paraíba do Sul. DZUFRJ-2046, 3 machos - São Fidélis, rio Paraíba do Sul. DZUFRJ-2044, 1 macho - São Fidélis, rio do Colégio (afluente do rio Paraíba do Sul). MNRJ-888, 1 macho e 1 fêmea - Araruama, lagoa de Juturnaíba. MNRJ-2286, 6 machos e 3 fêmeas - Maricá. MNRJ-2671, 3 fêmeas (2 ovadas) - Niterói, Jurujuba. MNRJ-2673, 1 macho - Rio de Janeiro, rio na encosta da Gávea. MNRJ-2297, 1 macho - Tinguá, represa da Boa Esperança. MNRJ-2676, 1 macho - Guaratiba, rio na estrada da Barra de Guaratiba. DZUFRJ-2116, 1 fêmea (ovada) - Mangaratiba, Fazenda Lindo Vale. DZUFRJ-3785, 2 machos - Mangaratiba, Fazenda Lindo Vale.
56 46 DZUFRJ-3790, 2 fêmeas - Mangaratiba, Fazenda Lindo Vale. DZUFRJ-2114, l macho - Mangaratiba, rio na praia do Saí. MNRJ- 1034, l macho - Angra dos Reis. MNRJ-1092, 1 macho e 2 fêmeas - Parati. Localidade tipo.- Jamaica (Sloane, 1758). O tipo foi depositado no British Museum, Londres, Inglaterra. Distribuição geográfica.- Desde a Flórida e Texas (EUA), até Santa Catarina (Brasil), incluindo as Antilhas. Observações.- Espécimens desta espécie são habitantes de água doce e salobra. São encontrados em trechos de rio com correnteza e também em remansos com fundo de areia e pedras. Preferem locais com fluxo constante de água (ESCOBAR, 1979). Parte do seu desenvolvimento (primeiros estágios) é realizado em água salgada e salobra (HOLTHUIS, 1980). De acordo com ESCOBAR, 1979 são encontrados em altitudes de até 200 metros. Macrobrachium olfersii (Wiegmann, 1836). Est. VIII (a-e) Astacus 987 Gronovius, 1764:23 1, pl. 17, fig. 1. Astacus Serratus Meuschen, 1781:9. Palaemon olfersii Wiegmann, 1836: 150.
57 47 Macrobrachíum olfersíí; Pearse, 1911: Holthuis, 1952:95, pi. 25, figs. a, - Villalobos, 1969: Gomes-Corrêa, 1977:53, est. XI, figs Williams, 1984:70, figs Abele & Kim 1989: Macrobrachíum bíraí Lobão, Melo & Fernandes, 1986:1. Caracterização.- Rostro curto, reto ou um pouco voltado para baixo e alcançando a extremidade distal do pedúnculo antenular; margem dorsal com 12 a 15 dentes dispostos regularmente, dos quais 4 ou 5 estão localizados sobre a carapaça, atrás da margem orbital; margem ventral com 3 a 4 dentes. Carapaça lisa. Abdome liso. Pleura do quinto somito abdominal com ângulo posterior reto ou ligeiramente agudo. Margem posterior do telso com espinhos internos mais longos que a sua extremidade aguda. Primeiro par pereópodes alcança a extremidade distal do escaf ocerito com 1/3 do comprimento do carpo. Dedos tão longos quanto a palma, ou um pouco mais curtos; carpo duas vezes o comprimento do própode; mero quase tão longo quanto o carpo. Segundo par de pereópodes do macho adulto muito desigual em forma e tamanho e com espinhos em todos os artículos. O maior ultrapassa a extremidade distal do escafocerito com o carpo inteiro e uma pequena parte do mero; dedos curtos deixando um espaço entre eles, quando fechados; cada margem cortante com um dente forte na porção proximal, dois dentes menores entre o dente grande e a base dos dedos e outros dentes menores dispostos regularmente até a extremidade distal do dedo; margens cortantes flanqueadas por pêlos duros. Palma bastante dilatada, sendo 1.5 a 2 vezes mais longa que larga, ligeiramente maior que o dátilo coberta com fortes espinhos e parcialmente coberta com densa pubescência. Carpo tão longo quanto o mero. Segundo pereópode menor ultrapassa a extremidade distal do escafocerito com parte do carpo; palma não dilatada; dedos
58 vez mais longos do que a palma e com espaço entre eles, quando fechados; cada margem cortante com apenas um dente em sua porção proximal e muitos pêlos longos e eretos. Terceiro par de pereópodes alcança a extremidade distal do escafocerito com a extremidade do dátilo; própode 2.5 a 2.8 vezes mais longo do que o dátilo, 1.5 a 2 vezes do que o carpo e menor que o mero. Quarto par de pereópodes com própode 2.5 vezes mais longo do que o dátilo, 1.5 a 2 vezes do que o carpo e quase tão longo quanto o mero. Quinto par de pereópodes 2.5 a 3.5 mais longo do que o dátilo, 1.5 a 2 vezes do que o carpo e tão longo quanto o mero. Tamanho.- O comprimento dos machos varia de 16 a 75 mm e o das fêmeas ovadas fica entre 28 e 54 mm. Os ovos são numerosos e pequenos. Material examinado: DZUFRJ-2042, 8 machos e 2 fêmeas - Itaperuna, rio Muriaé. MNRJ-3354, 11 machos e 6 fêmeas (3 ovadas) - Barra de Itabapoana. MZUSP-1913, 1 macho - rio Paraíba do Sul. MZUSP-12037, 3 machos - rio Paraíba do Sul. MNRJ-862, 1 macho - Itaocara, rio Paraíba do Sul. DZUFRJ-2047, 1 macho - Itaocara, rio Paraíba do Sul. DZUFRJ-2043, 5 machos e 8 fêmeas (6 ovadas) - São Fidélis, Cambiasca, rio Negro. DZUFRJ-2045, 1 macho - São Fidélis, rio Dois Rios (afluente do rio Paraíba do Sul). DZUFRJ-2048, 7 machos e 13 fêmeas (ovadas) - São Fidélis, rio Dois Rios ( afluente do rio Paraíba do Sul).
59 49 DZUFRJ-2049, 4 machos e 8 fêmeas (4 ovadas) - São Fidélis, rio Dois Rios ( afluente do rio Paraíba do Sul). DZUFRJ-2050, 1 fêmea - São Fidélis, rio Dois Rios (afluente do rio Paraíba do Sul). DZUFRJ-2053, 2 machos - São Sebastião do Alto, rio Grande (afluente do rio Dois Rios). DZUFRJ-2052, 3 machos e 2 fêmeas - São Sebastião do Alto, rio Negro. DZUFRJ-2051, 3 machos - São Sebastião do Alto, rio Grande (afluente do rio Dois Rios). MNRJ-10 machos e 14 fêmeas (7 ovadas) - Macaé, rio d'anta. MNRJ-2750, 4 machos e 2 fêmeas (ovadas) - rio das Ostras, riacho que desemboca na praia do Mar do Norte. MNRJ-2666, 1 macho e 1 fêmea - Barra de São João, rio São João. MNRJ-2727, 5 machos e 2 fêmeas - Nova Friburgo, rio Robelo. MNRJ-2575, 1 macho - Nova Friburgo, rio Ipitiuna (cruzando a estrada de Itaocara). MZUSP-5945, 1 macho e 1 fêmea (ovada) - Cachoeiras de Macacu, no Guapiaçu. MZUSP-5947, 1 macho - Cachoeiras de Macacu, rio Guapiaçu. MNRJ-4400, 1 macho - Rio Bonito, rio Bonito. DZUFRJ-1972, 5 machos e 7 fêmeas (3 ovadas) - Maricá, rio Fundo. DZUFRJ-2030, 1 fêmea - Maricá, lagoa de Maricá. DZUFRJ-2027, 1 macho e 2 fêmeas - Maricá, rio Fundo. DZUFRJ-3564, 3 machos e 6 fêmeas - Maricá, rio Bananal. DZUFRJ-3572, 3 machos - Maricá, rio Engenho Novo. DZUFRJ-2025, 21 machos e 7 fêmeas (3 ovadas) - Maricá, rio Engenho Novo. DZUFRJ-2029, 6 machos - Maricá, rio Engenho Novo.
60 50 DZUFRJ-2022, 14 machos e 29 fêmeas - Maricá, rio Bananal. MNRJ-2294, 1 fêmea - Maricá, serra da Tiririca, córrego dos Colibris. DZUFRJ-2031, 2 machos e 3 fêmeas - Magé, Santo Aleixo, rio Andorinhas. DZUFRJ-3563, 1 macho - Magé, rio Inhomirim, Raiz da Serra. MNRJ-2568, 1 macho - Rio de Janeiro, riacho que desemboca na praia de Copacabana. Col. em 03/12/1911. MNRJ-2570, 3 fêmeas - Rio de Janeiro, Fumas da Tijuca. Col. em 06/05/1962. MNRJ-2576, 1 macho - Rio de Janeiro, canal da Barra da Tijuca. Col. em 19/05/1969. MNRJ-2577, 1 macho - Rio de Janeiro, Jacarepa gu á, rio do Anil. Coletado em 01/11/1942. MNRJ-3428, 2 fêmeas - Rio de Janeiro, lagoa de Jacarepaguá. MNRJ-3375, 9 machos e 5 fêmeas - Barra de Guaratiba, riacho na estrada da Ilha. MNRJ-972, 1 fêmea - lta gu aí, rio Guandu. MNRJ-4373, 2 machos e 1 fêmea (ovada) - Ita gu aí, rio Guandu. MNRJ-1 macho - Coroa Grande. MNRJ-2593, 2 machos e 1 fêmea - Itacuruçá, rio Itin gu açu. MNRJ-2725, 1 fêmea (ovada) - Itacuruçá, Á gu as Lindas. MNRJ-2566, 2 machos - Ilha da Marambaia. MNRJ-2591, 2 machos - Ilha da Marambaia. MNRJ-2590, 4 machos e 4 fêmeas - Ilha da Marambaia. DZUFRJ-2119, 1 macho e 7 fêmeas (1 ovada) - Mangaratiba, rio na praia do Saí. DZUFRJ-2121, 3 machos e 4 fêmeas - Mangaratiba, rio Grande. MNRJ-2572, 36 machos e 40 fêmeas - Mangaratiba, Conceição de Jacareí. MNRJ-3368, 2 machos - Angra dos Reis, Ilha Grande.
61 51 DZUFRJ-2060, 3 machos e 2 fêmeas -Angra dos Reis, Ilha Grande. DZUFRJ-2037, 21 machos e 38 fêmeas (1 ovada) - Resende, Fazenda Santa Mônica. DZUFRJ-2062, 1 macho - Angra dos Reis, no Grande (estrada entre Mangaratiba e Lídice ). DZUFRJ-2065, 6 machos e 5 fêmeas - Angra dos Reis, rio Bracuí. MNRJ-3353, 6 machos e 6 fêmeas -Angra dos Reis, rio Mambucaba. DZUFRJ-2107, 1 macho e 3 fêmeas - Parati, rio São Roque. DZUFRJ-2105, 6 machos 1 fêmea -Parati, rio São Roque. DZUFRJ-2104, 2 machos e 1 fêmea (ovada) - Parati, rio São Roque. DZUFRJ-2093, 2 machos - Parati, rio São Roque. DZUFRJ-2095, 10 machos e 2 fêmeas - Parati, rio São Roque. DZUFRJ-2072, 1 macho - Parati, rio São Roque. MNRJ-2771, 5 machos - Parati, rio São Roque. DZUFRJ-2108, 1 macho - Parati, rio Barra Grande. DZUFRJ-2111, 18 machos e 17 fêmeas (7 ovadas) - Parati, rio Parati-Mirim. DZUFRJ-2090, 9 machos e 15 fêmeas - Parati, rio Parati-Mirim. DZUFRJ-2094, 3 machos e 5 fêmeas (3 ovadas) - Parati, rio Parati-Mirim. DZUFRJ-2079, 1 macho - Parati, rio Parati-Mirim. DZUFRJ-2078, 1 macho e 3 fêmeas - Parati, rio Parati-Mirim. DZUFRJ-2070, 2 fêmeas - Parati, rio Parati-Mirim. DZUFRJ-2088, 1 macho - Parati, rio Parati-Mirim. DZUFRJ-2086, 2 machos e 3 fêmeas (2 ovadas) - Parati, rio Parati-Mirim. MNRJ-4165, 3 machos e 5 fêmeas (ovadas) - Parati, rio Parati-Mirim. MNRJ-4 161, 1 fêmea - Parati, afluente na margem direita do rio Parati- Mirim. MNRJ-3330, 1 fêmea - Parati, rio Parati-Mirim.
62 Localidade tipo.- Costa brasileira. O tipo foi depositado no Zoologisch Musewn, Berlim, Alemanha. 52 (Brasil). Distribuição Geográfica.- Carolina do Norte (EUA) até Santa Catarina Observações.- Espécimens desta espécie são encontrados em águas doces e salobra, em fundo de areia e pedras. Segundo ESCOBAR (1979) podem ser encontrados em altitude de até 250 metros em trechos de rio com fundo de areia e lodo e perto de estuário, se escondendo sob troncos, folhas e pedras. Macrobrachium olfersii é bastante semelhante a M diguetti (BOUVIER, 1895) que ocorre do lado do Pacífico, podendo ser diferenciado desta, por possuir a palma do segundo par de pereópodes maior, dilatada e com maior número de cerdas longas (GOMES-CORRÊA, 1977). Macrobrachium potiuna (Müller, 1880). Est. IX ( a-d) Palaemon Potiuna Müller, 1880: 152. Macrobrachium potiuna Luederwaldt, 19 19:430 - Holthuis, 1952:76, pl. 18, figs. a d - Gomes-Corrêa, 1977:69, est. XVI, figs Bond & Buckup, 1983: Jalihal, Sankolli & Shenoy, 1993:373.
63 53 Caracterização.- Rostro curto e reto não ultrapassando a extremidade distal do pedúnculo antenular; margem dorsal com 5 a 1 O dentes distribuídos regulannente, sendo 1 ou 2 dentes localizados sobre a carapaça, atrás da margem orbital; margem ventral com 1 ou 2 dentes. Machos adultos com carapaça áspera na região anterolateral, devido à presença de pequenos espinhos. Abdome liso. Pleura do quinto somito abdominal com a extremidade um pouco aguda. Telso com destacada extremidade aguda na margem posterior e, também com espinhos internos longos, ultrapassando a citada extremidade. Primeiro par de pereópodes ultrapassa a extremidade distal do escafocerito com o própode inteiro e uma pequena parte do carpo. Dedos tão longos quanto a palma; carpo um pouco menos que 2 vezes mais longo do qtie o própode. segundo par de pereópodes do macho adulto igual em forma e um pouco desigual no tamanho, ultrapassando o escafocerito com parte do carpo. Dedos quase tão longos quanto a palma, cruzando quando fechados e deixando um espaço entre eles; margens cortantes com 2 dentes grandes na metade proximal do dátilo e um dente grande no dedo fixo que se encaixa entre os 2 dentes do dátilo. O dedo fixo ainda apresenta uma fileira de 4 pequenos dentes na porção proximal. Própode 2.5 vezes mais longo do que o carpo e o mero. Espinhos presentes em todos os artículos. Terceiro par de pereópodes alcançando a extremidade distal do escafocerito com a extremidade do dátilo, própode 3 vezes mais longo do que o dátilo, 2 vezes do que o carpo e ligeiramente menor que o mero. Quarto par de pereópodes 3.5 vezes tão longo quanto o dátilo, 2 vezes do que o carpo e tão longo quanto o mero. Quinto par de pereópodes com própode 4 vezes mais longo do que o dátilo, 2 vezes do que o carpo e um pouco maior que o mero. Tamanho.- O comprimento dos machos varia de 16 a 60 mm e o das fêmeas ovadas fica entre 28 e 50 mm. Os ovos são grandes e em pouca quantidade.
64 54 Material examinado: MNRJ-2636, 16 machos e 29 fêmeas (5 ovadas) - Santa Maria Madalena, serra Maria Madalena, córrego Tamanduá. DZUFRJ-2144, 3 fêmeas - Macaé, lagoa Cabiúnas. MNRJ-2633, 1 fêmea - Silva Jardim. DZUFRJ-3427, 135 machos e 122 fêmeas (3 ovadas) - Nova Friburgo, represa da CEDAE. MNRJ-6 machos e 8 fêmeas (4 ovadas) - Rio Bonito. MNRJ-2302, 2 machos - Teresópolis, Buracada Canoas. MNRJ-2635, 21 machos e 25 fêmeas (2 ovadas) - Petrópolis, estrada de Petrópolis. DZUFRJ-2035, 2 machos e 1 fêmea - Três Rios, rio Bemposta (afluente do rio Calçado). MNRJ-2631, 22 machos e 26 fêmeas - Areal, riacho da Fazenda da Lagoa. MNRJ-2621, 8 machos e 18 fêmeas - Areal, rio Preto. MNRJ-2620, 5 machos e 2 fêmeas - Duque de Caxias, rio Paô. Coletado em 27/06/1941. MNRJ- 2626, 1 fêmea (ovada) - Rio de Janeiro, riacho que desembocava na praia de Copacabana. Col. em Dez./ MNRJ-1703, 1 macho - Rio de Janeiro, Jacarepaguá, rio Camorim (próximo à represa Camorim). MNRJ-2628, 3 machos e 1 fêmea (ovada) - Rio de Janeiro, Jacarepaguá, rio Fundo. MNRJ-2639, 2 machos e 2 fêmeas - Rio de Janeiro, Jacarepaguá, Covanca. MNRJ-2623, 3 machos e 2 fêmeas (1 ovada) - Rio de Janeiro, Jacarepaguá, rio V argem Grande.
65 55 DZUFRJ-2132, 3 fêmeas (ovadas) - Miguel Pereira, cachoeira do rio Santa Branca (BR-125). DZUFRJ-2135, 2 machos e 1 fêmea - Miguel Pereira, rio Santa Branca (ponte do Livramento). DZUFRJ-2136, 3 machos - Miguel Pereira, no Santa Branca (ponte do Livramento). MNRJ-2634, 6 machos e 13 fêmeas - Nova Iguaçu, Reserva Biológica do Tinguá. MNRJ-3333, 1 fêmea - Vassouras. DZUFRJ-1992, 14 machos e 16 fêmeas (7 ovadas) - Campo Grande, represa do Mendanha. DZUFRJ-1991, 3 machos e 3 fêmeas (1 ovada) - Campo Grande, represa do Mendanha. DZUFRJ-2039, 13 machos - Campo Grande, represa do Mendanha. DZUFRJ-3425, 21 machos e 6 fêmeas - Campo Grande. MNRJ-2596, 7 machos - Campo Grande, rio Guandu Sena. MZUSP-9839, 1 macho - Itaguaí. DZUFRJ-2118, 2 machos e 13 fêmeas - Mangaratiba, serra do Piloto. DZUFRJ-2124, 9 machos e 3 fêmeas - Lídice, afluente do rio da Guarda (estrada de Angra dos Reis). DZUFRJ-2037, 21 machos e 38 fêmeas (1 ovada) - Quatis. DZUFRJ-2063, 11 machos e 22 fêmeas - Angra dos Reis, Matutu ( entre Mangaratiba e Lídice ). MNRJ-2610, 2 machos e 2 fêmeas - Angra dos Reis. MNRJ-2637, 6 machos e 2 fêmeas (1 ovada) - Angra dos Reis, Japuíba. MNRJ-2619, 1 macho - Angra dos Reis, rio Ariró. MNRJ-2758, 5 machos e 5 fêmeas - Resende, rio Pirapetinga (afluente do rio Paraíba do Sul)
66 56 MNRJ-2620, 1 macho e 3 fêmeas - Resende, rio Paraíba do Sul. MNRJ-2616, 2 machos - Itatiaia, rio do Cemitério. DZUFRJ-2100, 3 machos e 4 fêmeas (1 ovada) - Parati, cachoeira da Penha (200 metros). Localidade tipo.- Rio Itajaí (afluente do rio Itajaiaçu). O autor não designou tipo. Não há nenhuma referência feita à coleção na qual foram depositados os espécimens examinados pelo autor. Sul Distribuição geográfica. - Desde o Espírito Santo até o Rio Grande do Observações.- Espécie estritamente de água doce. Espécimens desta espécie podem ser capturados em rios com correnteza moderada com fundo de pedras e restos de vegetais, normalmente junto à vegetação marginal. De acordo com COELHO & RAMOS-PORTO (1985), são encontrados desde poucos metros a aproximadamente 1200 metros de altitude. M potiuna é bastante confundido com M iheringi (Ortmann, 1897), com o qual está proximamente relacionado. M potiuna apresenta os espinhos internos da margem posterior do telso muito longos, ultrapassando a sua extremidade aguda, já em M iheringi os mesmos espinhos não são tão longos e apenas alcançam a extremidade posterior aguda do telso. Os dedos do segundo par de pereópodes do macho adulto de M potiuna deixam um espaço quando fechados enquanto em M iheringi, fecham por inteiro. Muitas vezes toma-se dificil a diferenciação entre estas espécies, pois quando jovens mostram-se extremamente próximas, quanto às características acima citadas.
67 57 Macrobrachium iheringi (Ortmann, 1897) Est. X ( a-d). Palaemon iheringi Ortmann, 1897:211, pl. 1, figs Palaemon Iheringi Von Ihering, 1897:423. Macrobrachium iheringi - Luedeiwaldt, 1919:430 - Sawaya, 1946:406, pl. 1, fig. 1, pl. 2, fig. 2 - Holthuis, 1950: :85, pl. 21, figs. a-d - Gomes-Corrêa, 1977:73, est. XVII, figs Coelho & Ramos-Porto, 1985:405. Caracterização.- Rostro curto, com a extremidade um pouco voltada para baixo e alcançando a extremidade distal do pedúnculo antenular; margem dorsal com 6 a 9 dentes, dos quais 1 ou 2 estão localizados sobre a carapaça, atrás da margem orbital; margem ventral com 1 ou 2 dentes. Carapaça pode ter pequenos espinhos na sua região anterolateral. Abdome liso. Pleura do quinto somito abdominal terminando em um distinto pequeno ponto. Telso com margem posterior terminando em uma extremidade aguda, com espinhos posteriores internos alcançando esta extremidade. Primeiro par de pereópodes ultrapassa o escafocerito com o própode inteiro e uma parte do carpo. Dedos tão longos quanto a palma; carpo 1.5 vez mais longo do que o própode. Segundo par de pereópodes igual na forma e desigual no tamanho e com espinhos em todos os artículos. O pereópode maior ultrapassa o escafocerito com o própode inteiro e uma parte do carpo, quando esticado. Dedos não deixam espaço entre eles quando fechados; margens cortantes com 4 a 6 dentes na porção proximal; dátilo tão longo quanto a palma; própode 2 vezes mais longo do que o carpo. Terceiro par de pereópodes ultrapassa o escafocerito com uma parte do
68 58 dátilo, própode 2 a 2.5 vezes mais longo do que o dátilo, 2 vezes do que o carpo e tão longo quanto o mero. Quarto e quinto pares de pereópodes com própode 2.5 vezes mais longo do que o dátilo, 2 vezes do que o carpo e tão longo quanto o mero. Tamanho.- O comprimento dos machos examinados varia de 15 a 62 mm, e o das fêmeas ovadas fica entre 26 e 50 mm. Os ovos são grandes e em pouca quantidade. Material examinado: MNRJ-2652, 1 macho e 1 fêmea - Porciúncula, Fazenda Cachoeira. MNRJ-2648, 1 fêmea - Bom Jesus de Itabapoana, afluente do rio Itabapoana. DZUFRJ-2143, 3 fêmeas - Cantagalo, córrego da Areia. DZUFRJ-2032, 16 machos e 8 fêmeas - Bom Jardim, rio Grande. MNRJ-2757, 1 macho - Cachoeiras de Macacu, rio Rabelo. MNRJ-2653, 57 machos e 35 fêmeas - Magé, riacho no Distrito de Tocaia. MNRJ-2645, 1 macho - Rio de Janeiro, Jacarepaguá, lagoa de Camorim. MNRJ-2650, 4 fêmeas - Rio de Janeiro, Jacarepaguá. MNRJ-2647, 2 machos e 5 fêmeas - Nova Iguaçu, serra do Tinguá. MNRJ-3379, 2 machos e 2 fêmeas - Nova Iguaçu, Bela Vista, serra do Tinguá. MNRJ-2655, 84 machos e 120 fêmeas (15 ovadas) - Campo Grande, no Guandu Sena. MNRJ-969, 1 macho - Mangaratiba. MNRJ-2646, 1 fêmea - Itatiaia, Fazenda Serra, represa Itatiaia. MNRJ-2649, 5 machos e 1 fêmea - Itatiaia, rio Bonito.
69 Localidade tipo.- Rio Tietê, Estado de São Paulo, Brasil. Lectótipo e paralectótipos no Carnegie Musewn at Pittsburg, Pensilvannia. 59 Distribuição geográfica.- Bacias do Sudeste do Brasil. Observações. - classificada como espécie continental. Espécie estritamente de água doce sendo, por isso, Os indivíduos jovens não possuem os pequenos espinhos na região anterolateral da carapaça. Esta espécie é muito próxima de M potiuna sendo dela diferenciada pela comparação dos machos adultos, sendo muito di:ficil a identificação dos indivíduos jovens. Segundo COELHO & RAMOS-PORTO, 1985, são encontrados desde poucos metros até 1200 metros de altitude.
70 60 4. DISCUSSÃO 4.1. SISTEMÁTICA No presente trabalho foram examinados indivíduos de dez espécies, distribuídas em duas familias (Atyidae e Palaemonidae) e quatro gêneros (Atya, Potimirim, Palaemon e Macrobrachium). Dentre estas espécies M olfersii, foi a que mostrou um grande número de variações morfológicas. Ultimamente foram descritas duas espécies muito próximas à M olfersii: M holthuisi Genofre & Lobão, 1978 de São Sebastião, litoral norte de São Paulo, e M birai Lobão, Melo & Fernandes, 1986 para a Iegião de Cananéia, sul do Estado de São Paulo. LOBÃO et al, 1986 consideraram M birai tão próxima de M olfersii que a incluiram no grupo olfersii de VILLALOBOS (1969). As espécies que constituem o grupo olfersii são: M digueti (Bouvier, 1895), M faustinum (Saussure, 1857), M acantochirus (Villalobos, 1966), M crenulatum (Holthuis, 1950) e M hancocki (Holthuis, 1950). Para reunir estas espécies no grupo olfersii, Villalobos baseou-se em caracteres do segundo pereópode maior que possibilitaram considerá-las espécies afins. Estas espécies são bastante semelhantes à M olfersii quanto aos formatos do rostro, carapaça e telso (OSTROVSKI, FONSECA & SILVA FERREIRA, no prelo). As diferenças e semelhanças foram observadas nas
71 comparações das descrições e dos tipos de M birai e M holthuisi, e estão indicadas no quadro I. 61 M birai é semelhante à M olfersii nas características 3 ( a-b ), 5 e 6 (a), e M holthuisi, nas características 2, 4 e 8, sendo a primeira espécie muito mais próxima à M olfersii do que a segunda, conforme o exame de todo o material, incluindo os tipos, depositado na coleção do Museu de Zoologia da Universidade de São Paulo (M holthuisi 29 exemplares, e M birai 6 exemplares). Durante a análise detalhada de todo o material identificado como M olfersii, foi observado que as características 2, 4, 6 (a-c) e 7, expressas no quadro I, não são constantes, pois a grande maioria dos indivíduos de M olfersii examinados, apresentam características das três espécies e alguns ainda têm características intermediárias. Além disso, no exame detalhado do holótipo de M birai foi observado que a extremidade da 5 ª pleura ( característica 8) forma um ângulo reto como M olfersii e não é mais aguda como consta em LOBÃO et al, Diante da extrema semelhança entre M birai e M olfersii e da inconstância observada nas outras características que separam ambas as espécies, M birai foi considerada no presente trabalho como sinônima de M olfersii. As espécies M potiuna e M iheringi são muito próximas, sendo sua diferenciação muito dificil quando os exemplares são jovens, pois nesse estágio M potiuna assemelha-se a M iheringi. Deste modo os espécimens muito jovens foram colocados a nível de gênero (Macrobrachium sp ). Os caracteres que os diferenciam quando adultos são: espinhos posteriores do telso em M potiuna são mais longos que a extremidade do telso. Em M iheringi eles apenas alcançam esta extremidade; dedos do 2 º pereópode do macho adulto em M potiuna deixam um espaço quando fechados. Em M iheringi eles fecham por inteiro.
72 QUADRO I : Características que diferenciam M olfersii, M birai e M holthuisi (tirado de OSTROVSKI, FONSECA & SILVA-FERREIRA, no prelo) e) muito inchada 7. 2 pereópode o carpo inteiro se estende além o carpo e, às vezes um pouco do mero se metade do carpo se estende além do escafocerito estendem além do escaf ocerito do escafocerito (comprimento) 8. 5 segmento abdominal ex1remidade da pleura extremidade da pleura mais aguda extremidade da pleura formando formando ângulo reto ânj?:ulo reto 1 O'I N 1 Macho Adulto ; CARACTERISTICAS M. oi[!rsii Wie mann M. birai Lobão, Melo & Fernandes M. holthuisi Genofre & Lobão 1. Fórmula rostral 12-15, (4-5} 12-15, (3-5} 13-15, (3-5} Dentes rostrais regularmente distribuídos 1 dente mais distanciado dos demais rej?:ulannente distribuídos a) mero 4/5 do carpo a) mero 4/5 do carpo a) mero 9/10 do carpo pereópode b) 1/3 do carpo se estende além b) 1/3 do carpo se estende além do b) extremidade distal do carpo se do escafocerito escafocerito estende além do escafocerito 4. 2 pereópode dedo fixo com 3 dentes na dedo fixo com molar tricúspide na dedo fixo com 3 dentes na porção (margens cortantes) porção proximal ( o mais distal porção proximal seguido por um dente proximal ( o mais distal é maior) é maior) forte 2 pereópode (dedos) dedos fortemente cruzados, dedos curvados e com espaço entre eles dedos levemente curvados, sem deixando um espaço entre eles espaços entre eles a) com espinhos fortes, exceto a) com espinhos fortes, exceto na porção a) espinulada na porção mediana interna mediana interna onde mostra uma onde mostra uma densa densa pubescência 6. 2 º pereópode (palma) pubescência 5. b) 2 vezes tão longa quanto alta b) 2 vezes tão longa quanto alta b) 1.5 vez tão longa quanto alta e) lateralmente comprimida e) comprimida
73 63 Estas espécies além de muito semelhantes, possuem uma mesma área de ocorrência, o que também dificulta a sua diferenciação quando os exemplares são Jovens. P. potimirim é muito semelhante à P. mexicana, que ocorre no México, Costa Rica, Jamaica e Porto Rico. Estas duas espécies se reúnem em um grupo natural caracterizado pela presença de epipodito no coxopodito do quarto par de pereópodes (HOLTHUIS, 1955). P. potimirim tem o apêndice masculino mais estreito e as depressões entre os lobos, na margem posterior, mais profundas. Inicialmente, a outra espécie do gênero Potimirim, representada na região estudada, era P. brasiliana Villalobos, A partir de 1969, CHACE & HOBBS a incluíram na lista de sinonímias de P. glabra, devido à sua extrema semelhança com a referida espécie. MANNING & HOBBS (1977) ignoraram este argumento considerando ainda a espécie P. brasiliana. VILLALOBOS em 1982, relata o fato de estar realizando um estudo com a finalidade de provar a validade de P. brasiliana. Este estudo, segundo o mesmo autor, era baseado na variação de forma e proporções do apêndice masculino. Por não se ter conhecimento, na literatura, dos resultados finais deste estudo, a espécie P. brasiliana foi mantida na sinonímia de P. glabra DISTRIBUIÇÃO Os padrões distribucionais das espécies de camarões de água doce nas bacias hidrográficas do Rio de Janeiro, mostram-se bastante influenciados por
74 64 fatores ambientais, os quais atuam em todas as fases de desenvolvimento destes ammais. A concentração de cloretos na água (salinidade) representa importante fator na localização das espécies. De acordo com a dependência ou não de salinidade, os camarões são divididos em duas categorias ( como visto na introdução): espécies litorâneas - dependentes do estuário; Atya scabra, A. gabonensis, Potimirim glabra, P. potimirim, Palaemon (Palaemon) pandaliformis, Macrobrachium acanthurus, M carcinus e M olfersii; espécies continentais - estritamente de águas doces; Macrobrachium potiuna e M iheringi. Apesar de a espécie P. northropi ocorrer no Estado do Rio de Janeiro, ela não foi abordada no presente trabalho, por ser considerada uma espécie marinha. Segundo COELHO (1963a) P. northropi é uma espécie que freqüenta apenas os lugares onde a salinidade durante a preamar é igual a do mar, só diminuindo após grandes chuvas; já P. panda/iformis começa a ocorrer nos limites de penetração de P. northropi em diante, onde há variação do nível de água devido à ação das marés. A observação do diâmetro e quantidade de ovos, também permite identificar uma espécie como litorânea ou continental. As espécies litorâneas produzem ovos pequenos e numerosos, enquanto as espécies continentais produzem ovos grandes e em menor quantidade, o que sugere um encurtamento dos estágios larvares dependentes do estuário, fato que pode ser comprovado pela grande quantidade de vitelo presente em seus ovos (RODRIGUEZ, 1980). Segundo VILLALOBOS (1959), A. scabra passa as primeiras fases do seu desenvolvimento próximo da costa, para depois retomar rio acima e distribuir-se em áreas bastante
75 65 afastadas do estuário. Quanto à A. gabonensis, C. B. POWELL (apud HOBBS & HART, 1982) observou que larvas obtidas de fêmeas coletadas em água doce, morriam após cerca de 1 O a 12 dias, quando mantidas em água doce, mas com a adição de água salgada, sobreviviam até atingir um segundo estágio de desenvolvimento. Fatores ambientais como temperatura, oxigênio e declividade também são muito importantes na distribuição destes camarões, mas não foram medidos ao longo deste trabalho, pois grande parte dos exemplares estudados são de coleções. Apesar disso é possível relacioná-los à altitude e velocidade do rio, conforme citado em literatura. Como visto anteriormente, a declividade do terreno influencia diretamente na velocidade do fluxo de seus rios (correnteza), devido a maior energia potencial de suas águas. Quanto maior a correnteza no trecho de um rio, maior será a turbulência e mistura de suas águas e, consequentemente maior o nível de oxigênio dissolvido. A declividade de um rio determinará o seu gradiente ( está relacionado ao gradiente topográfico), ou seja, quanto mais acentuada for a inclinação de um terreno maior será o gradiente de seus rios. Esses fatores também vão determinar o tipo de leito do rio, sendo rochoso nas partes mais altas e arenoso ou lamoso nas partes mais baixas (próximas ao litoral), como resultado da deposição de sedimentos obtidos da erosão das partes mais altas. Entretanto, em alguns rios de pequeno porte, próximos ao litoral, seu fundo pode ser de cascalho. No litoral sul do Estado do Rio de Janeiro (Angra dos Reis, Parati) isto acontece. São rios que estão muito próximos a relevos de grande amplitude e possuem fluxos torrenciais em épocas de grandes chuvas, favorecendo o transporte de material grosseiro (J. MARQUES, com. pes. ). Assim sendo, as espécies litorâneas podem estar relacionadas a fundos de areia e lama ou de cascalho, já as espécies continentais estão mais relacionadas à fundo rochoso.
76 66 Como foi visto na introdução, as águas que vêm de áreas mais elevadas são mais frias. Assim sendo espécies encontradas em altitudes maiores, como M potiuna e M iheringi, parecem estar melhor adaptadas às condições descritas acima, tendo sido encontradas em altitudes de até 500 metros. Os limites de altitude estabelecidos por COELHO (1985) - de poucos metros a 1200 m - confirmam este dado. Segundo BOND & BUCKUP (1983), M potiuna é uma espécie!ótica, isto é, vive em águas continentais em movimento, precisando de elevados teores de saturação de oxigênio. M iheringi pertence à mesma área de ocorrência de M potiuna estando, portanto, adaptada às mesmas condições ambientais. No presente trabalho a espécie A. scabra foi encontrada desde poucos metros, em rios litorâneos, até aproximadamente 300 m de altitude em Natividade de Carangola local onde também foi encontrada A. gabonensis. Estas espécies são consideradas por HUNTE (1978) como espécies!óticas, estando perfeitamente adaptadas a ambientes com baixa altitude em rios rasos, de alto gradiente e com uma boa tolerância a temperaturas mais altas. De acordo com HOBBS & HART (1982) a distribuição destes camarões está restrita a trechos de rio de correnteza rápida a moderada e, portanto, com alto teor de oxigênio dissolvido. Como adaptação a este tipo de ambiente, estes animais possuem pereópodes fortes e cobertos com tubérculos escamiformes, permitindo que se agarrem ao fundo pedregoso, e impedindo que sejam arrastados pela correnteza (VILLALOBOS, 1982). Habitar massas d'água isoladas, representa uma desvantagem para os animais dulciaquícolas, já que isso limita seu potencial de dispersão. A solução para este problema está na retenção do estágio planctônico marinho, que mantém o
77 seu potencial de dispersão garantindo, assnn, a escolha do rio apropriado pela própria larva (HUNTE, 1978). 67 Mecanismos de dispersão podem aumentar a distribuição de algumas espécies pelo continente. Segundo VILLALOBOS (1982), existem espécies de Macrobrachium (M acanthurus e M carcinus) que podem sair do meio aquático para ultrapassar obstáculos que dificultem sua migração pela água, principalmente rio acima. M o/fersii também parece usar o mesmo artificio descrito anteriormente pois, como as espécies anteriormente citadas, apresenta uma distribuição bastante ampla dentro da área estudada, ocorrendo desde o litoral até 250 metros de altitude (de acordo com ESCOBAR, 1979) e foi encontrada no presente trabalho 1 indivíduo a 500 metros de altitude, no rio Grande ( entre Mangaratiba e Lídice ). De todas as espécies encontradas, M acanthurus foi a que se mostrou mais amplamente distribuída, tendo sido encontrada em todas as bacias hidrográficas. O fato de as larvas de Macrobrachium dependerem do estuário, permite sua dispersão pelo litoral por meio das correntes de maré. Indivíduos jovens podem associar-se às raízes de plantas macrófitas sendo arrastadas pela correnteza até o mar para depois alcançar outros rios. Já os indivíduos adultos podem migrar de uma bacia para outra, nas nascentes dos rios (VILLALOBOS, 1982). De acordo com HOBBS & HART (1982), espécies do gênero Atya, também são capazes de sair da água por pequenas distâncias, durante suas migrações rio acima, uma vez que sobem pequenos rios tributários até quase a cabeceira, transpondo pequenas cascatas. Presume-se que estas espécies sejam capazes de atravessar divisões baixas entre as cabeceiras próximas de dois rios. Indivíduos de Potimirim e jovens de Atya também podem ser encontrados entre as
78 68 raízes de plantas macrófitas, podendo ser levados pela correnteza para o mar, de lá sendo arrastados para outros lugares (VILLALOBOS, 1959). Segundo J. S. MARQUES (com. pess.), rios de uma bacia podem ser capturados por rios de outra bacia, ocorrendo em áreas onde a divisão é baixa entre as cabeceiras de rios de bacias diferentes como, por exemplo, nas áreas de cabeceiras das bacias do Paraíba do Sul e Tietê. Já, no Rio de Janeiro, isso é mais dificil de ocorrer, devido à sua grande área serrana, que impede comunicação entre as cabeceiras. De acordo com a mesma fonte, podem ocorrer comunicações entre bacias em suas áreas mais baixas, em períodos de enchentes. Estas comunicações podem representar uma possibilidade de migração de espécies de uma bacia para outra. De forma semelhante, a migração de indivíduos jovens por correntes de maré, através do litoral, também poderá levá-los a alcançar estuários de rios pertencentes à outras bacias próximas. Assim sendo, os mecanismos de dispersão citados mostram-se de grande importância na distribuição geográfica destas espécies de camarões, constituindo uma forma de compensação às dificuldades impostas pelo fato de habitarem massas d'água isoladas.
79 69 6. CONCLUSÕES No Estado do Rio de Janeiro foram encontradas dez espécies de camarões na rede fluvial, quatro representantes da família Atyidae e seis da familia Palaemonidae. Devido a estreita semelhança entre M birai e M olfersii pode-se sugerir que a primeira seja colocada na sinonímia da última. De acordo com a dependência do estuário, as espécies são classificadas como espécies litorâneas e espécies continentais. A distribuição das espécies nos tipos de água pode ser vista no quadro III. Palaemon (Palaeander) northropi, não foi abordado neste trabalho por ser uma espécie de água salgada, embora ocorra em parte da área estudada. Macrobrachium acanthurus foi a espécie de mais ampla distribuição, sendo encontrada em todas as bacias hidrográficas. M carcinus e M olfersii ocorrem nas mesmas bacias hidrográficas. As espécies da família Atyidae estão representadas em um número menor de bacias hidrográficas do Rio de Janeiro. A espécie Atya gabonensis é pela primeira vez citada para o Rio de Janeiro, tendo sido registrada somente na bacia do Paraíba do Sul.
80 70 M heterochirus (Wiegmann, 1836) não ocorre na área estudada, como afirmaram Holthuis (1952) e Gomes-Corrêa (1977). O exame dos espécimens listados revelou que pertencem à M carcinus.
81 71 QUADRO II: hidrográficas. Distribuição das espécies encontradas nas bacias BACIAS HIDROGRÁFICAS ESPÉCIES o 1 2 3e Atya 1;abonensis X A. scabra X X Potimirim glabra X X X P. potimirim X X X Palaemon (Palaemon) pandaliformis X X X X X Macrobrachium acanthurus X X X X X X X X M carcinus X X X X X X X M olfersii X X X X X X X M potiuna X X X X X X M iherin1;i X X X X X 9 X X X X X X X X Legenda: O - rio Itabapoana 1 - contribuinte à lagoa Feia 2 - rios Macaé, São João e Una 3 e 4 - contribuinte aos sistemas lagunares de Araruama, Saquarema, Maricá, Itaipu e Piratininga 5 - contribuinte à baía de Guanabara 6 - rio Paraíba do Sul 7 - contribuinte ao sistema lagunar de Jacarepaguá 8 - contribuinte à baía de Sepetiba 9 - contribuinte à baía de Ilha Grande
82 QUADRO III: Distribuição das espécies quanto ao tipo de água e altitude onde são encontradas TIPOS DE ÁGUAS ESPÉCIES DOCE SALOBRA ALTITUDE (metros) Atya scabra X X O a400 Potimirim glabra X X O a 100 Potimirim potimirim X X O a 100 Palaemon (Palaemon) pandal formis X X O a 100 Macrobrachium acanthurus X X O a 250 Macrobrachium carcinus X X O a 200 Macrobrachium o(fersii X X O a 100 Macrobrachium potiuna X até 1200 Macrobrachium iherin i X até J N Atya abonensis X X
83 73 REFERÊNCIAS BIBLIOGRÁFICAS ABELE, L. G., lntroductions of Two Freshwater Decapod Crustaceans (Hymenosomatidae and Atyidae) into Central and North America. Crustaceana, Leiden, 23(3): , figs A New Species of Freshwater Shrimp (Genus Atya) from the Pacific Drainage of Panama. Proc. biol. Soe. Wash., Washington D.C., 88(6): 51.:.58, figs. 1, 2. ABELE, L. G. & N. BLUM, Ecological Aspects of the Freshwater Decapod Crustaceans of the Pedas Archipelago, Panamá. Biotropica, 97(4): , figs ABELE, L. G. & W. KIM, An Illustrated Guide to the Marine Decapod Crustacea of Florida. Florida Department of Environmental Regulation, Technical Series, 8(4) 1: xvi pp The Decapod Crustaceans of the Panama Canal. Smithson. Contr. Zool., Washington, D.C., 482: 1-50, 18 figs., 3 tables. BATE, C. S., On a New Genus, with Four New Species of Freshwater Prawns. Proc. zool. Soe. Lond., London, : , pls BOND, G. & L. BUCKUP, O cultivo de Macrobrachium borel/i (Nobili, 1896) e de Macrobrachium potiuna (Müller, 1880) em laboratório. (Crustacea:Decapoda:Palaemonidae). Revta bras. Biol., Rio de Janeiro, 43(2):
84 74 BOWMAN T. E. & L. G. ABELE, Classification of the recent crustacea. ln: BLISS, D. E. Biology of Crustacea. New York. Academic Press. 1(1): BOUVIER, E. L., Sur les Palémons recuillis dans les eaux douces de la Basse - Californie par M digueti. Bull Mus. Hist. nat. Paris, Paris, 1: , figs Crevettes de la famille des Atyidés: Especes qui font partie des collectiom du Museum d'histoire Naturelle. Bull Mus. Hist. nat. Paris, Paris, 10: Observations nouvelles sur les crevettes de la famille Atyidés. Bull scient. Fr. Belg., 39: , figs Recherches sur la morphologie, les variations, la distribuition geographique des crevettes de la famille de Atyidés. Encycl. ent., Ser. A, 1: 1-370, 716 figs. BRUSCA, R. C. I. & G. J. BRUSCA, Invertebrates. Sunderland. Sinaur Associates,, pp BURKENROAD, M. D., The evolution of the Eucarida (Crustacea:Eumalacostraca) in relation to the fossil record. Tulane Stud. Geol. i(l): CALMAN, W. T., On the Classification of the Crustacea Malacostraca. Ann. Mag. nat. Hist., London, 1: CHACE, F. A. JR., The Shrimps of the Smithsonian - Bredin Caribbean Expedition - with a summary of the W est lndian Shallow - Water Species
85 (Crustacea - Decapoda - Natantia). Smithson. Contr. Zool., Washington, D.C., 98: x+ 179, figs. 75 CHACE, F. A., JR. & H. H. HOBBS, JR., The Freshwater and Terrestrial Decapoda Crustaceans of the West Indies with Special Reference to Dominica. Bull U.S. natn. Mus., Washington, D.C., 292: pp, 76 figs., 5 pl. COELHO, P. A., Observações preliminares sobre a biologia e a pesca dos camarões do gênero Macrobrachium Bate, 1868 (Decapoda, Palaemonidae) no Estado de Pernambuco, Brasil. Trabs. Inst. Oceanogr. Univ. Pernambuco, Recife,_l: a. Algumas observações sobre a biologia e a ecologia dos camarões Pa/aemon northropi e P. panda/iformis no Estado de Pernambuco (Decapoda:Palaemonidae). Trab. Inst. Oceanogr. Univ Pernambuco., Recife, 5/6: COELHO, P. A. & M. A. RAMOS, A contribuição e a distribuição da fauna de decápodos do litoral leste da América do Sul entre as latitudes de 5 º N e 39 S. Trabs. Oceangr. Univ. Fed. Pernambuco, Recife, li: COELHO, P. A. & M. RAMOS-PORTO, Camarões de água doce do Brasil: Distribuição Geográfica. Revta bras. Zool., São Paulo, i(6): CHRISTOFOLETTI, A.,1981. Geomorfologia Fluvial. Vol. 1, O Canal Fluvial. Edgard Blücher, p. DANA, J. D., Conspectus of the Crustacea of the United States Exploring Expedition under command of Capt. Charles Wilkes, U. S. N. Proc. Acad. nat. Sei. Philad., Philadelphia,, :
86 76 ESCOBAR, J. G., Carideos (Palaemonidae y Atyidae) en los rios de la Region de Santa Marta. An. lnst. Inv. Mar. Punta Betin. Santa Marta. Colombia, 11: F ABRICIUS, J. C., Su1wlementum entomologiae systematicae, pp Hafniae. FAUSTO-FILHO, J., Crustáceos Decápodos de Valor Comercial ou Utilizados como Alimento no Nordeste Brasileiro. Bolm Soe. cearense Agron., Fortaleza,.2_: FELGENHAUER, B. E. & L. G. ABELE, Feeding structures of two Atyid shrimps, with comments on Caridean phylogeny. J. Crustacean Biol., Boston, (3): FONSECA, K. M. L.; M. e. OSTROVSKI & T. e. G. SILVA-FERREIRA, Ocorrência de Atya gabonensis Giebel, 1875 (Crustacea:Decapoda:Atyidae) na bacia do rio Paraíba do Sul, Estado do Rio de Janeiro. Nauplius, Rio Grande,. : GENOFRE, G. C. & V. L. LOBÃO, Macrobrachium holthuisi sp. n. a new species of shrimp (Decapoda, Macrura). Crustaceana, Leiden, 35(3): GIEBEL, C. G., Atya gabonensis, neuer Krebs aus Gabon. Zeitschr. ges. Naturwiss., 45: GOMES-CORRÊA, M. M., Palaemonídeos do Brasil (Crustacea, Decapoda, Natantia). Dissertação de Mestrado, Universidade Federal do Rio de Janeiro, 164 pp, 28 ests.
87 77 GRONOVIUS, T., Zoophylacium Gronovianun Fasciculus secundus exhibens enumerationem Insectorum, quae in museo suo adservat, examini subjecit, systematice disposuit atque descripsit. Part 2, Pages , 17 plates. Leiden: Sumptibus Auctoris. HAAN, W. de, Crustacea. ln: P.F. de SIEBOLD. Fauna Japonica sive descriptio animalium quae in itinere per Japoniam, jussu et auspiciis superiorum qui summum in India Batavia Imperium tenent, suscepto, annis collegit, notis, observationibus et adumbrationibus illustrativ: IX-XVI, I-XXXI, VII-XVII, 243 pp, pls. A-Q, A. Arnz (Lugdunum Batavorum). HART, C. W., JR., Toe freshwater shrimps (Atyidae and Palaemonidae) of Jamaica. W. I.; with a dicussion of their relation to the ancient geography of the Westem Caribbean area. Proc. Acad. nat. Sei. Philad., Philadelphia, : ,, New Freshwater Shrimp Records for Tobago, West Indies, With a Summary of Records for the Lesser Antilles (Atyidae and Palaemonidae). Proc. biol. Soe. Wash., Washington, D.C., 93(3): , 1 fig. HEDGPETH, J. W., The North American Species of Macrobrachium (River Shrimp). Texas J. Sei., l: 28-38, figs HOBBS, H. H., JR., Atya gabonensis (Decapoda, Atyidae) in the Westem Hemisphere. Crustaceana, Leiden, 38(1): 111. HOBBS, H. H., JR. & C. W. HART, JR., The Shrimp Genus Atya (Decapoda, Atyidae). Smithson. Contr. Zool., Washington, D.C., 364: 1-143, 53 figs.
88 78 HOLTHUIS, L. B.,1950. Toe Palaemonidae collected by the Siboga and Snelius Expeditions with remarks on other species. I - Sub-family Palaemoninae. Siboga-Exped. Leiden, Leiden, 39a 9, 10: 1-267, figs Toe Caridean Crustacea of Tropical West Africa. Atlantide Rep., i: 7-187, figs A general revision of the Palaemonidae (Crustacea, Decapoda, Natantia) of the Americas. II. Toe subfamily Palaemoninae. 0cc. Pap. Allan Hancock Fdn, Los Angeles, 12: 1-396, pls On a collection of decapod crustacea from the Republic of El Salvador (Central America). Zool. Verh., Leiden, 23: 1-43, 15 text. figs., 2 pls.,, 1954a. Observaciones sobre los crustáceos decápodos de la República El Salvador. Comun Inst. Trop. Invest. Cient. de El Salvador, J(4): The recent genera of Caridean and Stenopodidean shrimps ( Class Crustacea, Order Decapoda, Supersection N atantia) with keys for their determination. Zool. Verh., Leiden, 26: 1-157, 104 text-figs Toe crustacea decapoda of Suriname (Dutch. Guiana). Zool. Verh., Leiden, 44: 1-296, pls , Toe R/V Pillsbury Deep-Sea Biological Expedition to the Gulf of Guinea, , 11: Toe Freshwater Shrimps of the Island of Annobon, West Africa. Stud. trop. Oceanogr., 4 (1): , 5 figs.
89 FAO Species Catalogue, 1: Shrimps and Prawns of the world: An Annotated Catalogue of Species of Interest to Fisheries. F.A. O. Fish. Biol. Synopses, Roma, 125(1): Fresh-water Shrimps of the Family Atyidae (Crustacea:Crustacea) from Westem Colombia. (3): , figs J. Crustacea Biol., Boston,, Toe Decapod Crustacea of Brokopondo lake, Surinam. Nauplius, Rio Grande, 1: HUNTE, W., The distribution of freshwater shrimps (Atyidae and Palaemonidae) in Jamaica. J. Linn. Soe., London, 64: , 7 figs. IHERING, H. von, Os camarões de água doce do Brasil. Revta Mus. paul., São Paulo, i: JALIHAL, D. R., K. N. SANKOLLI & S. SHENOY, Evolution of development pattems and the process of freshwaterization in the prawn genus Macrobrachium Bate, 1868 (Decapoda, Palaemonidae ). Crustaceana, Leiden, 65 (3): KINGSLEY, J. S., Notes on the North American Caridea in the Museum of the Peabody Acaderny of Science at Salem. Mass. Proc. Acad. nat. Sei. Philad., Philadelphia: KOELBEL, C., Carcinologisches. Sber. Akad. Wiss. Wien, 90(1): , 3 plates. LATREILLE, P. A., Genera crustaceorum et insectorum secundum ordinem naturalem in families disposita. 1: Paris.
90 80 LEACH, W. E., Atys scaber. Trans. Lin. Soe. Lond., London,,2: 345.,, Atya. ln: Encyclopedia Britannica, Supplement to the fourth, fifth and sixty editions, 1(1): 421, plate 21. Edimburg: Archibald Constable and Company [Reprinted 1824]. LEMOS DE CASTRO, A., Sobre os crustáceos referidos por Marcgrave em sua "Historia Naturalis Brasiliae" (1648). Archos Mus. nac. Rio de J. Rio de Janeiro, 52: 37-51, est LINNAEUS, C., Systema Naturae per Regna tria Naturae, secundum Classes, Ordines, Genera, Species, Cum Characteribus, Differentiss, Synonymis, locis. (ed. 10) 1, iii+824 pp. Estocolmo. LOBÃO, V. L., G. A. S. MELO & V. M. FERNANDES, Redescrição de Macrobrachium. birai, Lobão, Melo & Fernandes e de Macrobrachium petronioi, Melo, Lobão & Fernandes (Crustacea:Decapoda), Palemonídeos da região sul do Estado de São Paulo, Brasil. Bolm. lnst. Pesca, São Paulo, 15 (1): LUEDERWALDT, H., Lista dos crustáceos superiores (Thoracostraca) do Museu Paulista que foram encontrados no Estado de São Paulo. Revta Mus. paul., São Paulo, U: MANNING, R. B. & HOBBS Jr., H. H., Decapoda. ln: S. H. Hulbert, ed., Biota Aguatica de Sudamerica Austral, : San Diego State University, San Diego, California. MARCGRAF, G., Historiae Rerum Naturalium Brasiliae Libri Octo: Quorum tres priores 1 de Plantis. Ouartus de Piscibus Quintus de A vibus, Sextus de Quadrupedibus et Serpentibus.
91 81 MEUS CHEN, F. C., lndex, continens nomina generica specierum propria, trivialia ut et synonyma. ln: L. T. Gronovius, editor, Zoolophylacium Gronovianum, exhibens Anirnalia Ouadrupeda, Amphibia, Pisces, Isecta, Vermes, Mollusca, Testacea, et Zoophyta, quae in museo suo adsevavit, examini subjecit. systematice disposuit atque descripsit, 19 unnumbered pages. Leiden: T. Haak & Co., S. and J. Luchtmans. MILNE-EDW ARDS, A., Revision des Crustacés Macroures de la famille de Atyoidées. Annls. Soe. Ent. Fr., Paris, series 4, 1_: , pl. 3. MONOD, TH., L'industrie des pêches au Cameroun: Mission Monod ( ) Cameroun. Volume 1, unnumbered pages, 96 figures, 25 plates. Paris: Société d'editions Geographiques, Maritimes et Colonaise Sur quelques Crustacés de l'afrique Occidentale (Liste des Décapodes mauritaniens et des Xanthidés ouest-africains). Bull. Com. Étud. hist. scient. Afr. Occid. fr., 15(2-3): , 26 figs Crevettes et crabes de la côte occidentale d'afrique. ln. Réunion de spécialistes C. S. A. sur les Crustacés. Mémoires l'lnstitut Fondamental d'afrique Noire, 77: , 26 pls Sur quelques Crustacés Décapodes africains (Sahel-Soudan). Bull. Mus. nat. Hist. Nat, Paris, 3 ª Sér. Zool, 500: Décapodes. ln: J. R. Durand and C. Lévêque, editors, Flore et faune aquatiques de l'afrique Sahélo-Soudanienne, Paris, 1: MOREIRA, C., Crustáceos do Brasil. Contribuição para o conhecimento da fauna brasileira. Archos Mus. nac. Rio de!., Rio de Janeiro., li:
92 82 MÜLLER F., Palaemon potiuna. Ein Beispel abgekürzter Verwandlung. Zool. Anz., J: ,, Atyoida potimirim, Eine schlam fressende Susswassergameele. Kosmos, : , 20 figs. OLIVEIRA, L. P. H., Verificação da existência de Atya scabra Leach, Camarão d'água doce da família Atyidae, Crustacea, no nordeste do Brasil. Mems. Inst. Oswaldo Cruz, Rio de Janeiro, 43(1): ORTMANN, A. E., Die Decapoden-Krebse des Strassburger Museums, I. Theil. Zool. Jahrb. Syst. Geogr. Biol. Thiere, : , pi. 1, 2.,, A study of the Systematic and Geographical Distribution of the Decapod Family Atyidae Kingsley. Proc. Acad. nat. Sei. Philad., Philadelphia, 46 (1984): ,, Os camarões de água doce da América do Sul. Família Atyidae Kingsley. Revta Mus. paul., São Paulo, i: OSTROVSKI, M. C., K. M. L. FONSECA & T. C. G. SILVA-FERREIRA, no prelo. Macrobrachium denticulatum sp. n. a new species of shrimp from the São Francisco basin, northeastem Brazil (Decapoda, Palaemonidae ). Crustaceana, Leiden. PEARSE, A. S., Report on the Crustacea collected by the University of Michigan - Walker Expedition in the State of Vera Cruz, Mexico. Rep. Michigan Acad. Sei., Michigan, li: , figs. 1-4.
93 83 RAFINESQUE, C. S., Precis des Decouvertes et Travaux somiologisches. Palenne: Royale Typographie Militarie. (Lithoprint reproduction, wakefield, Massachussetts: Toe Murray Printing Co., 1948), 55 pp.. RANDALL, J. W., Catalogue of the Crustacea brought by Thomas Nutall and J. K. Towsend, from the west Coast of North America and the Sandwiches Islands, with descriptions of such species as are apparently new among which are included several species of different localities, previosly existing in the collection of the Academy. J. Acad. Nat. Sei, Philadelphia, -ª.: , lams RATHBUN, M. J., Toe Decapod Crustacea of Jamaica. Ann. Inst. Jamaica, 1(1): 45pp Toe Brachyura and Macrura of Porto Rico. Buli U. S:. Fish. Comm., Washington, i: Pls. I-II. RODRIGUEZ, G., Los crustaceos decapodos de Venezuela. Inst. venez. Invest. Cien., Caracas, 450 p. RODRIGUEZ, G., Decapoda. ln: S.H. HURLBERT, G. Rodriguez and N.D. SANTOS, (ed). Aquatic Biota of Tropical South America, Part 1: Arthropods, : San Diego State University, San Diego, California. SAUSSURE, H. de, Diagnoses de quelques Crustacés nouveaux de l'amérique tropicale. Revue Mag. Zool., Paris, (2) _2: SAWAYA, M. P., Sobre alguns camarões d'água doce do Brasil. Archos Zool. Est. S. Paulo, São Paulo, 11: , pls
94 FALCÃO, M. M., Bacias Hidrográficas do Estado do Rio de Janeiro (SERLA). Secretaria de Estado de Obras e Meio Ambiente, Rio de Janeiro. SLOANE, H., A Voyage to the Island Madera, Barbados, Nieves, St. Christophers and Jamaica, with the Natural Histmy of the Herbs and Trees, Four-footed Beasts, Fishers, Birds, Insects, reptiles & C of the Last of Those Islands to Wich is Prefix'd an Introduction, Werein is an Account of the Inhabitants, Air, Water, Diseases, Trade, & C of that Place, with some Relations Concerning the Neighbouring America. 2 vols. xvii+499 pp, xi+274 pls. SMALLEY, A. E., Toe genus Potimirim in Central America (Crustacea: Atyidae). Revta biol. Trop., Costa Rica, 11(2): SMITH, S. 1., List of the Crustacea collected by J. A. McNiel in Central America. Rep. Peabody Acad. Sei., (1869); STIMPSON, W., Notes on North American Crustacea in the Museum of the Smithsonian Institution. III. Ann. Lyceum nat. Hist., New York, 10: THOMPSON, D. W., A catalogue of Crustacea and of Pycnogonida contained in the Museum of University College, Dundee. Univ. St. Andrews, Dundee, 56 p. VILLALOBOS, A. F., Decap.). México, 70 p, 22 pls. Estudio morfológico de la Atya scabra (Crust. Tesis, Universidad Nacional Autónoma de México, Cidade do., Contribución al conocimiento de los Atyidae de Mexico (Crustacea, Decapoda). II. Estudio de algunas especies de Potimirim (=Ortmannia) con descripción de una especie de Brasil. An. Inst. Biol. Univ. Méx., México, 30:
95 85, Problemas de especiacion en America de un grupo de Palaemonidae del gênero Macrobrachium. F.A.O. Fisheries Repotl, Roma, 57 (3): Decapoda. ln: S. H. > HURLBERT and A. VILLALOBOS - FIGUEROA, editors, Aquatic Biota of Mexico, : (San Diego State University, San Diego, California). WEBER, F., Systematicam ill. viii+ 171 p. Nomenclator entomologicus Secundum Entomologiam Fabrici adjectis speciebus recens defectis et varietatibus. WIEGMANN, A. F. A., Reschreibung einiger neuer crustaceen des Berliner. Museums aus México und Brasilien. Arch Naturgesch., 2.(1): WILLIAMS, A. B., Marine Decapod Crustaceans of the Carolinas. Fish Bull Wildl. Serv., Washington, D.C., 65(1): 1-298, figs., Shrimps, Lobsters and Crabs of the Atlantic Coast of the Eastem United States Maine to Florida. Washington, D.C. Smithsonian Inst. Press, xviii+550 pp, 380 text figs
96 MAPAS: DISTRIBUIÇÃO DAS ESPÉCIES NO ESTADO DO RIO DE JANEIRO Escala aproximada - 1 :
97 Fonte: SERLA, 1985 (Superintendência Estadual de Rios e Lagos) - Estado do Rio de Janeiro - Secretaria de Estado de Obras e Meio Ambiente. Mapa l Unidades Hidrográficas* O. rio ltabapoana l. contribuinte à lagoa Feia 2. rios Macaé, São João e Una 3 e 4. contribuinte aos sistemas lagunares de Araruama, Saquarema, Marie Itaipu e Piratininga 5. contribuinte à baía de Guanabara 6. rio Paraíba do Sul 7. contribuinte ao sistema lagunar de Jacarepaguá 8. contribuinte à bafa de Sepetiba 9. contribuinte à baía de Ilha Grande,f e ' l. r, (.1._;1. J e. ( ' e. ( e>

98 "-' \ o --..,,,, - -r 1 30' 22 2 ;' ' l---' ffi] 21 t MINAS GERAIS 1 cy /' OIC E A NO 10' 44 º 1n 4 º 30 ' '
99 Mapa Atya gabonensis Atya scabra Potimirim glabra Potimirim potimirim \ ( (, 1 l ( ) ( (', e ( 1 ' ( ( )

100 º 30 ' t 30' ,r-----t j 30'
101 '1' e (, (, ( \ / j < ( ) ( ( e: ( \( r ( l. q / ( ( Mapa 3 5. Palaemon (Palaemon) pandaliformis 6. Macrobrachium acanthurus

102 ' cy, ' ' ' 23 I ' ; '
103 Mapa 4 1. Macrobrachium carcinus 8. Macrobrachium olfersii r, \... ( ( (, ( (, ( (,f l \ (.. (

104 , cy O' ' ' ' ' '
105 Mapa 5 9. Macrobrachium potiuna 10. Macrobrachium iheringi ( ( ( (


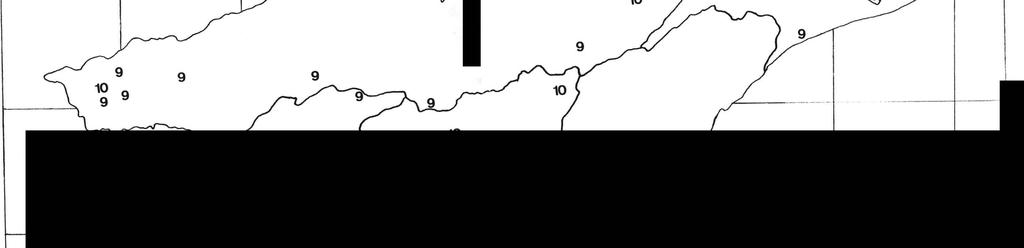
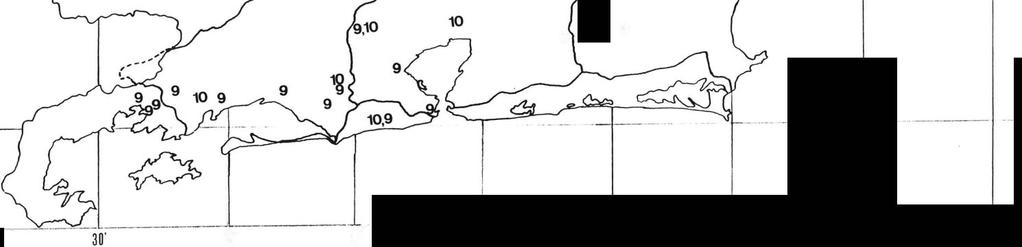
106 _ ' í 30' _l ' 43 30'
107 V v
108 ESTAMPAS
109 Atya gabonensis (macho) a.- Vista dorsal da região cefálica; b.- Vista lateral da região cefálica; e.- Vista lateral do terceiro pereópode; d.- Superficie flexora da porção distal do terceiro pereópode; e.- Apêndices masculino e interno; f- Vista lateral da carena preanal; g.- Vista lateral da segunda à quinta pleura abdominal; h.- Vista dorsal do telso. Obs.: Escalas marcadas em 1 mm. V < >Fonte: Hobbs & Hart, 1982.



110 a r r 1 a, b, e, g e d f g h...- "
111 Aiya scabra (macho) a.- Vista dorsal da região cefálica; b.- Vista lateral da região cefálica; c.- Vista lateral do terceiro pereópode; d.- Superficie flexora da porção distal do terceiro pereópode; e.- Apêndices masculino e interno; f- Vista lateral da carena preanal; g.- Vista lateral da segunda à quinta pleura abdominal; h.- Vista dorsal do telso. Obs.: Escalas marcadas em 1 mm. ( ) Fonte: Hobbs & Hart, 1982.



112 J.. - _/ r a, b, e, g d p <1 e, f g h h
113 ESTAMPA III Potimirim glabra (macho) a.- Vista lateral da carapaça; b.- Vista lateral do segundo pereópode; c.- Apêndices masculino e interno; d.- Vista dorsal do telso e urópodes.

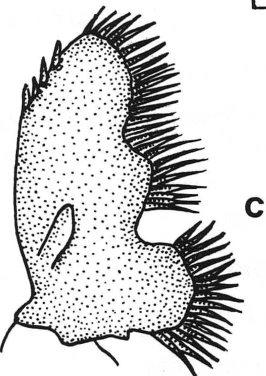
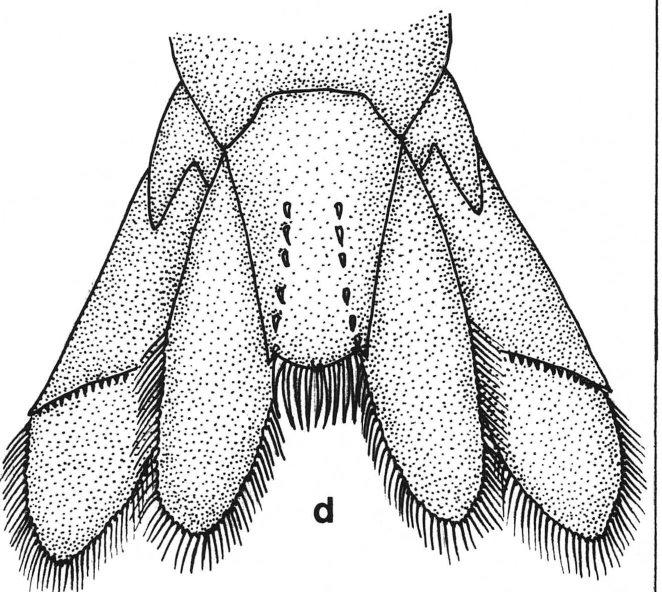
114 a ' : : , mm mm
115 ESTAMPA IV Potimirim potimirim (macho) a.- Vista lateral da carapaça; b.- Vista lateral do segundo pereópode; c. - Apêndices masculino e interno; d.- Vista dorsal do telso e urópodes..,
mfa: ':.-:./.\:?")


116 1 mm llfi\r,- ;:,;,.;.;:.,;; :. >..._.,. _'.-i >'?/-??>)mfa: ':.-:./.\:?:.:-- ::/.:-\/://: º ::;::-:.::::;', = :-.::-::...: :::-;:::i :..: '.....' : : ' : : :{:: : :. :. a d.,: :-: : :. 0.5 mm 2.5 mm
117 ESTAMPA V Palaemon (Palaemon) pandaliformis (macho) a. - Vista lateral da região cefálica; b.- Vista lateral do segundo pereópode; c.- Vista dorsal do telso e urópodes; d.- Extremidade posterior do telso
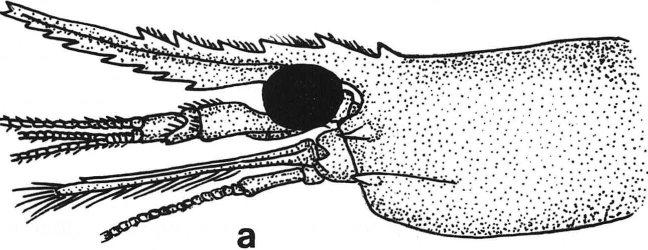
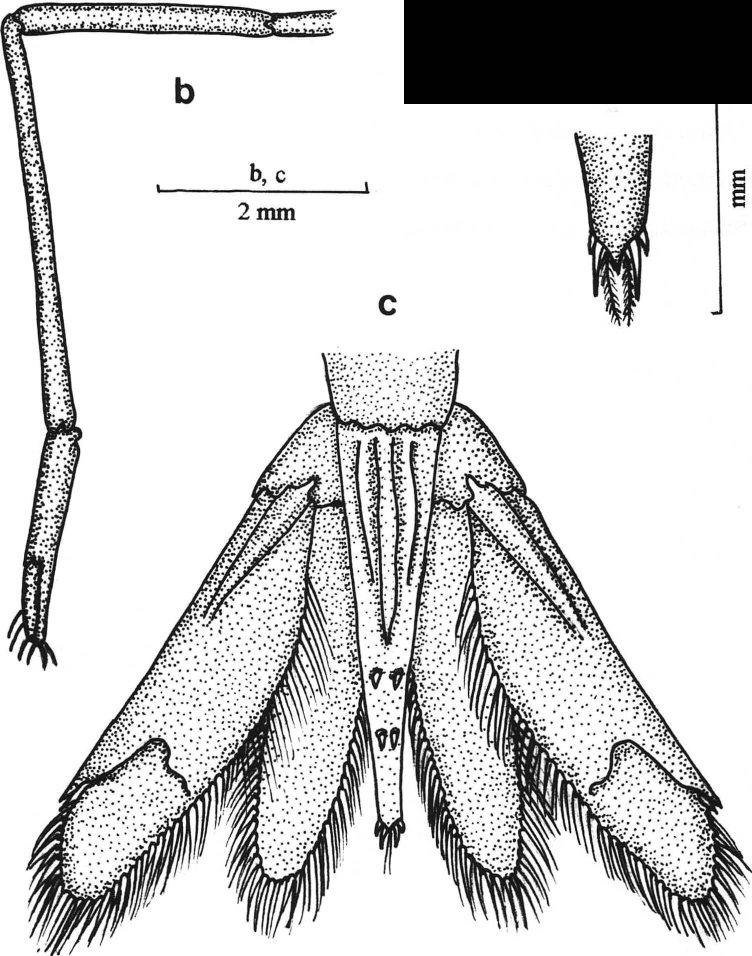
118 2mm d -
119 L- ESTAMPA VI.Macrobrachium acanthurus (macho) a.- Vista lateral da região cefálica; b. - Vista lateral do segundo pereópode; e.- Dedos do segundo pereópodes (sem pêlos); d.- Extremidade posterior do telso; e.- Vista dorsal do telso e urópodes.

120 e '. d a, b, e 5mm d 1 mm e 5mm
121 ,.. ESTAMPA VII Nfacrobrachium carcinus (macho) a.- Vista lateral da região cefálica; b.- Vista lateral do segundo pereópode; e.- Vista dorsal do telso e urópodes; d.- Extremidade posterior do telso.


122 d a '.... :,..... '.-.: _.._ :. _ :... 2mm 5 mm a, e snun e
123 ESTAMPA VIII lvf acrobrachium olfersii (macho) a.- Vista lateral da região cefálica; b.- Vista lateral do segundo pereópode maior; e.- Vista lateral do segundo pereópode menor; d.- Vista dorsal do telso e urópodes; e.- Extremidade posterior do telso.

124 5mm b, e 5mm
125 ESTAMPA IX Afacrobrachium potiuna (macho) u a.- Vista lateral da região cefálica; b.- Vista lateral do segundo pereópode; e.- Vista dorsal do telso e urópodes; d.- Extremidade posterior do telso.

126 a, b 5mm d 1 mm
127 ESTAMPA X Afacrobrachium iheringi (macho) a. - Vista lateral da região cefálica; b.- Vista lateral do segundo pereópode: V e.- Vista dorsal do telso e urópodes; d.- Extremidade posterior do telso.


128 :.... a, b 5 mm d e 2mm 1 mm
Galatheoidea Chirostylidae Uroptycbus uncifer (A. Milne Edwards, 1880)
") Anomura Galatheoidea Chirostylidae -- Uroptycbus uncifer (A. Milne Edwards, 1880) Diagnose : Esta espécie se diferencia essencialmente de U. nitidus pelas dimensões de seus quelípodos que são mais largos
Anomura Galatheoidea Chirostylidae -- Uroptycbus uncifer (A. Milne Edwards, 1880) Diagnose : Esta espécie se diferencia essencialmente de U. nitidus pelas dimensões de seus quelípodos que são mais largos
Munida pusilla Benedict, 1902
 t~* Anomura Galatheoidea Galatheidae *- Munida pusilla Benedict, 1902 Diagnose : Espécie de tamanho pequeno. Carapaça com bordas arqueadas e pouco mais longa do que larga, convexa e com margem anterior
t~* Anomura Galatheoidea Galatheidae *- Munida pusilla Benedict, 1902 Diagnose : Espécie de tamanho pequeno. Carapaça com bordas arqueadas e pouco mais longa do que larga, convexa e com margem anterior
Uma nova espécie de anfípode cavern ícola do Brasil Hyalel/a caeca sp.n. (Amphipoda, Hyalellidae).
.") REVISTA BRASILEIRA DE ZOOLOGIA Revta bras. Zoo!., 6 ( 1 ) : 4 9 - S S ls/rv/1989 Uma nova espécie de anfípode cavern ícola do Brasil Hyalel/a caeca sp.n. (Amphipoda, Hyalellidae). Vânia F. Goulart C. Pereira
REVISTA BRASILEIRA DE ZOOLOGIA Revta bras. Zoo!., 6 ( 1 ) : 4 9 - S S ls/rv/1989 Uma nova espécie de anfípode cavern ícola do Brasil Hyalel/a caeca sp.n. (Amphipoda, Hyalellidae). Vânia F. Goulart C. Pereira
GUATUBESIA, NOVO GÊNERO DE GONYLEPTIDAE
 Rev. bras. Ent. 22í2): 99-104 11.1X.1978 OPERA OPILIOLOGICA VARIA. XV. GUATUBESIA, NOVO GÊNERO DE GONYLEPTIDAE Helia E.M. Soares Guatubesia clarae, n. gen., n. sp. (Opiliones, Gonyleptidae, Gonyleptinae),
Rev. bras. Ent. 22í2): 99-104 11.1X.1978 OPERA OPILIOLOGICA VARIA. XV. GUATUBESIA, NOVO GÊNERO DE GONYLEPTIDAE Helia E.M. Soares Guatubesia clarae, n. gen., n. sp. (Opiliones, Gonyleptidae, Gonyleptinae),
Camarões de águas continentais (Crustacea, Caridea) da Bacia do Atlântico oriental paranaense, com chave de identificação tabular
 da Bacia do Atlântico oriental paranaense, com chave de identificação tabular") Acta Biol. Par., Curitiba, 38 (1-2): 11-34. 2009. 11 Camarões de águas continentais (Crustacea, Caridea) da Bacia do Atlântico oriental paranaense, com chave de identificação tabular Freshwater shrimps
Acta Biol. Par., Curitiba, 38 (1-2): 11-34. 2009. 11 Camarões de águas continentais (Crustacea, Caridea) da Bacia do Atlântico oriental paranaense, com chave de identificação tabular Freshwater shrimps
CLASSE MALACOSTRACA SUBCLASSE EUMALACOSTRACA SUPERORDEM EUCARIDA
 CLASSE MALACOSTRACA SUBCLASSE EUMALACOSTRACA SUPERORDEM EUCARIDA carapaça fundida com todos os segmentos da cabeça e toráx - cefalotórax. télson com ramos caudais 0 a 3 pares de maxilípodos brânquias na
CLASSE MALACOSTRACA SUBCLASSE EUMALACOSTRACA SUPERORDEM EUCARIDA carapaça fundida com todos os segmentos da cabeça e toráx - cefalotórax. télson com ramos caudais 0 a 3 pares de maxilípodos brânquias na
Roteiro Aula Prática Biologia dos Crustáceos. Prof.Dr. Alvaro Luiz Diogo Reigada
 Roteiro Aula Prática Biologia dos Crustáceos Prof.Dr. Alvaro Luiz Diogo Reigada Classe Malacostraca Ordem Stomatopoda O segundo par de antenas é trirreme, abdome espesso e robusto. Carapaça curta recobrindo
Roteiro Aula Prática Biologia dos Crustáceos Prof.Dr. Alvaro Luiz Diogo Reigada Classe Malacostraca Ordem Stomatopoda O segundo par de antenas é trirreme, abdome espesso e robusto. Carapaça curta recobrindo
Oi pessoal dos 8 os anos, estão com saudade das atividades escolares? Pois bem, precisamos dar seqüência aos estudos e, na volta às aulas, no dia 17,
 Oi pessoal dos 8 os anos, estão com saudade das atividades escolares? Pois bem, precisamos dar seqüência aos estudos e, na volta às aulas, no dia 17, algumas atividades devem estar prontas. Mas antes de
Oi pessoal dos 8 os anos, estão com saudade das atividades escolares? Pois bem, precisamos dar seqüência aos estudos e, na volta às aulas, no dia 17, algumas atividades devem estar prontas. Mas antes de
l P ^ ^ C DESCRIÇÃO DE TRÊS ESPÉCIES NOVAS DE MAJIDAE DO BRASIL (DECAPODA: BRACHYURA)
") CDU 595.384.2 rir-j ü' r -- % li l P ^ ^ C - /1 ^ DESCRIÇÃO DE TRÊS ESPÉCIES NOVAS DE MAJIDAE DO BRASIL (DECAPODA: BRACHYURA) PETRÔNIO ALVES COELHO Laboratório de Ciências do Mar da Universidade Federal
CDU 595.384.2 rir-j ü' r -- % li l P ^ ^ C - /1 ^ DESCRIÇÃO DE TRÊS ESPÉCIES NOVAS DE MAJIDAE DO BRASIL (DECAPODA: BRACHYURA) PETRÔNIO ALVES COELHO Laboratório de Ciências do Mar da Universidade Federal
CRUSTACEA. classes. Malacostraca. Maxillopoda. Branchiopoda. Cephalocarida. Remipedia. Baseado e modificado de Brusca et al.
 CRUSTACEA classes 2 Malacostraca Maxillopoda 1 Branchiopoda Cephalocarida Remipedia 1 Larva náuplio Desenvolvimento pós-embrionário anamórfico Metameria homônoma Apêndices birremes tipo filopódio (plesio)
CRUSTACEA classes 2 Malacostraca Maxillopoda 1 Branchiopoda Cephalocarida Remipedia 1 Larva náuplio Desenvolvimento pós-embrionário anamórfico Metameria homônoma Apêndices birremes tipo filopódio (plesio)
Tipos de Ecossistemas Aquáticos
 UNIVERSIDADE FEDERAL DO RIO GRANDE LICENCIATURA EM CIÊNCIAS INTERDISCIPLINA FENÔMENOS DA NATUREZA II Tipos de Ecossistemas Aquáticos Dr. Cleber Palma Silva Dra. Edélti Faria Albertoni Lab. Limnologia Para
UNIVERSIDADE FEDERAL DO RIO GRANDE LICENCIATURA EM CIÊNCIAS INTERDISCIPLINA FENÔMENOS DA NATUREZA II Tipos de Ecossistemas Aquáticos Dr. Cleber Palma Silva Dra. Edélti Faria Albertoni Lab. Limnologia Para
SISTEMÁTICA DE Rhabdepyris KIEFFER (HYMENOPTERA, BETHYLIDAE) AMAZÔNICOS COM ANTENAS PECTINADAS
 AMAZÔNICOS COM ANTENAS PECTINADAS") SISTEMÁTICA DE Rhabdepyris KIEFFER (HYMENOPTERA, BETHYLIDAE) AMAZÔNICOS COM ANTENAS PECTINADAS Adriano da S. MAGESKY 1, Celso Oliveira AZEVEDO 1 RESUMO - São descritas e ilustradas Rhabdepyris opistolatus
SISTEMÁTICA DE Rhabdepyris KIEFFER (HYMENOPTERA, BETHYLIDAE) AMAZÔNICOS COM ANTENAS PECTINADAS Adriano da S. MAGESKY 1, Celso Oliveira AZEVEDO 1 RESUMO - São descritas e ilustradas Rhabdepyris opistolatus
OS PALAEMONIDAE DE ÁGUAS CONTINENTAIS DO BRASIL MERIDIONAL (CRUSTACEA, DECAPODA)
") yím1 Rev. BrasU. Biol., 49(4):883-896 Novembro, 1989 - Rio de Janeiro, RJ OS PALAEMONIDAE DE ÁGUAS CONTINENTAIS DO BRASIL MERIDIONAL (CRUSTACEA, DECAPODA) GEORGINA BOND-BUCKUP e LUDWIG BUCKUP 1 2 Departamento
yím1 Rev. BrasU. Biol., 49(4):883-896 Novembro, 1989 - Rio de Janeiro, RJ OS PALAEMONIDAE DE ÁGUAS CONTINENTAIS DO BRASIL MERIDIONAL (CRUSTACEA, DECAPODA) GEORGINA BOND-BUCKUP e LUDWIG BUCKUP 1 2 Departamento
MARCUSIAXIUS LEMOSCASTROI, g.n., sp.n., NOVA OCORRÊNCIA DA FAMÍLIA AXIIDAE (CRUSTACEA, DECAPODA, TH ALASSINIDE A) NO BRASIL
 NO BRASIL") /? ri//, i/0 > n -i - ff Boi. Zool. e Bioh Mar.. N.S.,»..76», pp. 553-566, São Paulo, W7-i dwstacbà LlBRAf SIIIITHSOHXAS I*-i. RETUBM TO V^' MARCUSIAXIUS LEMOSCASTROI, g.n., sp.n., NOVA OCORRÊNCIA DA FAMÍLIA
/? ri//, i/0 > n -i - ff Boi. Zool. e Bioh Mar.. N.S.,»..76», pp. 553-566, São Paulo, W7-i dwstacbà LlBRAf SIIIITHSOHXAS I*-i. RETUBM TO V^' MARCUSIAXIUS LEMOSCASTROI, g.n., sp.n., NOVA OCORRÊNCIA DA FAMÍLIA
OWESj m, Iftr ^ ^ ^ ^ ^ REDESCRIÇÃO DE "LEPIDOPA VENUSTA" (STIMPSON) E SUA OCORRÊNCIA NO LITORAL SUL DO BRASIL (Decapoda, Albuneidae) i
 E SUA OCORRÊNCIA NO LITORAL SUL DO BRASIL (Decapoda, Albuneidae) i") OWESj m, Iftr ^ ^ ^ ^ ^ IgflBi! m i 4 w ^ msnr M M CmióíACEA Rev. Brasil. Biol., 25 (1) : 97-103 Maio, 1965 - Rio de Janeiro, GB REDESCRIÇÃO DE "LEPIDOPA VENUSTA" (STIMPSON) E SUA OCORRÊNCIA NO LITORAL
OWESj m, Iftr ^ ^ ^ ^ ^ IgflBi! m i 4 w ^ msnr M M CmióíACEA Rev. Brasil. Biol., 25 (1) : 97-103 Maio, 1965 - Rio de Janeiro, GB REDESCRIÇÃO DE "LEPIDOPA VENUSTA" (STIMPSON) E SUA OCORRÊNCIA NO LITORAL
DESCRIÇÃO DE UMA ESPÉCIE NOVA DO GÊNERO "LEPIDOPA" STIMPSON, E SUA OCORRÊNCIA NO LITORAL BRASILEIRO (Decapoda, Albuneidae) 1
 1") Vs/ifK comjou^ev.fe oj autkor ^ Rev. Brasil. Biol., 28 (1) : 77-86 Abril, 1968 Rio de Janeiro, GB DESCRIÇÃO DE UMA ESPÉCIE NOVA DO GÊNERO "LEPIDOPA" STIMPSON, E SUA OCORRÊNCIA NO LITORAL BRASILEIRO (Decapoda,
Vs/ifK comjou^ev.fe oj autkor ^ Rev. Brasil. Biol., 28 (1) : 77-86 Abril, 1968 Rio de Janeiro, GB DESCRIÇÃO DE UMA ESPÉCIE NOVA DO GÊNERO "LEPIDOPA" STIMPSON, E SUA OCORRÊNCIA NO LITORAL BRASILEIRO (Decapoda,
Crustacea (L. crusta = concha)
") Subfilo Crustacea Crustacea (L. crusta = concha) - Caranguejos, siris, camarões, lagostas, krill; - Marinho água doce e terrestre; - Mais abundantes, diversificados e maior distribuição oceanos; - Todas
Subfilo Crustacea Crustacea (L. crusta = concha) - Caranguejos, siris, camarões, lagostas, krill; - Marinho água doce e terrestre; - Mais abundantes, diversificados e maior distribuição oceanos; - Todas
TAXONOMIA E PADRÕES DISTRIBUCIONAIS DOS CAMARÕES MARINHOS E ESTUARINOS DA FAMÍLIA PALAEMONIDAE (CRUSTACEA: DECAPODA: CARIDEA) DO LITORAL BRASILEIRO
 DO LITORAL BRASILEIRO") UNIVERSIDADE FEDERAL DO RIO GRANDE PÓS-GRADUAÇÃO EM OCEANOGRAFIA BIOLÓGICA TAXONOMIA E PADRÕES DISTRIBUCIONAIS DOS CAMARÕES MARINHOS E ESTUARINOS DA FAMÍLIA PALAEMONIDAE (CRUSTACEA: DECAPODA: CARIDEA)
UNIVERSIDADE FEDERAL DO RIO GRANDE PÓS-GRADUAÇÃO EM OCEANOGRAFIA BIOLÓGICA TAXONOMIA E PADRÕES DISTRIBUCIONAIS DOS CAMARÕES MARINHOS E ESTUARINOS DA FAMÍLIA PALAEMONIDAE (CRUSTACEA: DECAPODA: CARIDEA)
UM NOVO GÊNERO E DOIS ALÓTIPOS DE "GONY- LEPTIDAE" (OPILIONES) 1
 1") Rev. Brasil. Biol., 5(3) :339-34 3 Setembro, 19-15 Rio de Janeiro, D. F. UM NOVO GÊNERO E DOIS ALÓTIPOS DE "GONY- LEPTIDAE" (OPILIONES) 1 B. A. M, SOARES e HÉLIA ELLER MONTEIRO SOARE S Departamento de
Rev. Brasil. Biol., 5(3) :339-34 3 Setembro, 19-15 Rio de Janeiro, D. F. UM NOVO GÊNERO E DOIS ALÓTIPOS DE "GONY- LEPTIDAE" (OPILIONES) 1 B. A. M, SOARES e HÉLIA ELLER MONTEIRO SOARE S Departamento de
Universidade de São Paulo Instituto de Física de São Carlos Licenciatura em Ciências Exatas Biologia
 Universidade de São Paulo Instituto de Física de São Carlos Licenciatura em Ciências Exatas Biologia 2 2016 Aula Prática 7: ARTHROPODA UNIRAMIA: MYRIAPODA E INSECTA MYRIAPODA Os miriápodos possuem cabeça
Universidade de São Paulo Instituto de Física de São Carlos Licenciatura em Ciências Exatas Biologia 2 2016 Aula Prática 7: ARTHROPODA UNIRAMIA: MYRIAPODA E INSECTA MYRIAPODA Os miriápodos possuem cabeça
 '-vvj
'-vvj BOLETIM DO MUSEU NACIONAL NOVA SÉRIE RIO DE JANEIRO - BRASIL
 BOLETIM DO MUSEU NACIONAL NOVA SÉRIE RIO DE JANEIRO - BRASIL ISSN 0080-312X ZOOLOGIA N o 505 25 DE JULHO DE 2003 DUAS ESPÉCIES DO GÊNERO ACANTHOCARPUS STIMPSON, 1871 (CRUSTACEA, CALAPPIDAE) PARA A COSTA
BOLETIM DO MUSEU NACIONAL NOVA SÉRIE RIO DE JANEIRO - BRASIL ISSN 0080-312X ZOOLOGIA N o 505 25 DE JULHO DE 2003 DUAS ESPÉCIES DO GÊNERO ACANTHOCARPUS STIMPSON, 1871 (CRUSTACEA, CALAPPIDAE) PARA A COSTA
As terras submersas e a água no globo terrestre
 As terras submersas e a água no globo terrestre CONTINENTES Blocos continentais que resultaram da divisão do continente único: Pangeia. Quatro grandes massas continentais: Continente Tríplipe (África,
As terras submersas e a água no globo terrestre CONTINENTES Blocos continentais que resultaram da divisão do continente único: Pangeia. Quatro grandes massas continentais: Continente Tríplipe (África,
19) Leporinus friderici (Bloch, 1794)
 Leporinus friderici (Bloch, 1794)") 144 Ovos e larvas de peixes de água doce... 19) Leporinus friderici (Bloch, 1794) Nomes comuns: Aracu-comum, aracu-cabeça-gorda, aracu, aracu-branco, piava e piau. Distribuição geográfica: Bacias do Prata,
144 Ovos e larvas de peixes de água doce... 19) Leporinus friderici (Bloch, 1794) Nomes comuns: Aracu-comum, aracu-cabeça-gorda, aracu, aracu-branco, piava e piau. Distribuição geográfica: Bacias do Prata,
Manoel Luciano Aviz Quadros*, Cristiana Maciel, Sandra Bastos & Iracilda Sampaio
 REPRODUÇÃO DO CAMARÃO CANELA - MACROBRACHIUM ACANTHURUS EM CONDIÇÕES CONTROLADAS DE LABORATÓRIO E MONTAGEM DE UM ATLAS PARA IDENTIFICAÇÃO DE ESTÁGIOS LARVAIS Manoel Luciano Aviz Quadros*, Cristiana Maciel,
REPRODUÇÃO DO CAMARÃO CANELA - MACROBRACHIUM ACANTHURUS EM CONDIÇÕES CONTROLADAS DE LABORATÓRIO E MONTAGEM DE UM ATLAS PARA IDENTIFICAÇÃO DE ESTÁGIOS LARVAIS Manoel Luciano Aviz Quadros*, Cristiana Maciel,
30) Prochilodus lineatus (Valenciennes, 1836)
 Prochilodus lineatus (Valenciennes, 1836)") 198 Ovos e larvas de peixes de água doce... 30) Prochilodus lineatus (Valenciennes, 1836) Nomes comuns: Curimba, corimba, curimbatá e corimbatá. Distribuição geográfica: Bacia do Prata e do rio Paraíba
198 Ovos e larvas de peixes de água doce... 30) Prochilodus lineatus (Valenciennes, 1836) Nomes comuns: Curimba, corimba, curimbatá e corimbatá. Distribuição geográfica: Bacia do Prata e do rio Paraíba
Revta bras. Ent. 32(2): OPERA OPILIOLOGICA VARIA. XXI. (OPILIONES, GONYLEPTIDAE) ABSTRACT
: OPERA OPILIOLOGICA VARIA. XXI. (OPILIONES, GONYLEPTIDAE) ABSTRACT") Revta bras. Ent. 32(2):233-237. 26.09.1988 8 OPERA OPILIOLOGICA VARIA. XXI. (OPILIONES, GONYLEPTIDAE) Helia E.M.Soares1 Benedicto A.M. Soares* ABSTRACT Stefanesia avramae, gen. n., sp.n., described from
Revta bras. Ent. 32(2):233-237. 26.09.1988 8 OPERA OPILIOLOGICA VARIA. XXI. (OPILIONES, GONYLEPTIDAE) Helia E.M.Soares1 Benedicto A.M. Soares* ABSTRACT Stefanesia avramae, gen. n., sp.n., described from
ARQUIVOS DO MUSEU BOCAGE
 Vol. VI 1976 ARQUIVOS DO MUSEU BOCAGE (2." Série) nofas e suplementos n. 27 SOBRE A PRESENÇA EM PORTUGAL DE MUNIDA IRIS RUTLLANTI ZARIQUIEY ALVAREZ, 1952 (DECAPODA, ANOMURA, GALATHEIDAE) ANA M. NEVES (*)
Vol. VI 1976 ARQUIVOS DO MUSEU BOCAGE (2." Série) nofas e suplementos n. 27 SOBRE A PRESENÇA EM PORTUGAL DE MUNIDA IRIS RUTLLANTI ZARIQUIEY ALVAREZ, 1952 (DECAPODA, ANOMURA, GALATHEIDAE) ANA M. NEVES (*)
DEIPARTAMENTO DE ZOOLOGIA. DOIS NOVOS CiÊNEROS E TRÊS NOVAS ESPÉCIES DE OPILIOES BRASILEIROS (*) POR INTRODUÇÁO
 POR INTRODUÇÁO") Vol. V, N." 26 - pp. 243-250 PAPÉIS AVULSOS DO DEIPARTAMENTO DE ZOOLOGIA SECRETARIA DA AGRICULTURA - S. PAULO - BRASIL DOIS NOVOS CiÊNEROS E TRÊS NOVAS ESPÉCIES DE OPILIOES BRASILEIROS (*) POR INTRODUÇÁO
Vol. V, N." 26 - pp. 243-250 PAPÉIS AVULSOS DO DEIPARTAMENTO DE ZOOLOGIA SECRETARIA DA AGRICULTURA - S. PAULO - BRASIL DOIS NOVOS CiÊNEROS E TRÊS NOVAS ESPÉCIES DE OPILIOES BRASILEIROS (*) POR INTRODUÇÁO
Relação entre os grande grupos de artrópodes
 Relação entre os grande grupos de artrópodes Chelicerata Myiriapoda Crustacea Hexapoda Crustacea Pancrustacea Mandibulata Pancrustacea ou Tetraconata Evidências Morfologia do olho composto Formação sistema
Relação entre os grande grupos de artrópodes Chelicerata Myiriapoda Crustacea Hexapoda Crustacea Pancrustacea Mandibulata Pancrustacea ou Tetraconata Evidências Morfologia do olho composto Formação sistema
55) Hoplosternum littorale (Hancock, 1828)
 Hoplosternum littorale (Hancock, 1828)") Ordem Siluriformes - H. littorale 313 55) Hoplosternum littorale (Hancock, 1828) Nomes comuns: Caborja, tamboatá, camboatá e cascudinho. Distribuição geográfica: América do Sul a leste dos Andes e norte
Ordem Siluriformes - H. littorale 313 55) Hoplosternum littorale (Hancock, 1828) Nomes comuns: Caborja, tamboatá, camboatá e cascudinho. Distribuição geográfica: América do Sul a leste dos Andes e norte
31 OCORRÊNCIA DE LITHADIA ROTUNDATA A. MILNE EDWARDS, 1880 NO LITORAL BRASILEIRO (CRUSTACEA: BRACHYURA: LEUCOSIIDAE).
.") 31 OCORRÊNCIA DE LITHADIA ROTUNDATA A. MILNE EDWARDS, 1880 NO LITORAL BRASILEIRO (CRUSTACEA: BRACHYURA: LEUCOSIIDAE). MARIA FERNANDA ABRANTES TORRES (1) GUSTAVO AUGUSTO SCHMIDT DE MELO (2 ) 1 - Departamento
31 OCORRÊNCIA DE LITHADIA ROTUNDATA A. MILNE EDWARDS, 1880 NO LITORAL BRASILEIRO (CRUSTACEA: BRACHYURA: LEUCOSIIDAE). MARIA FERNANDA ABRANTES TORRES (1) GUSTAVO AUGUSTO SCHMIDT DE MELO (2 ) 1 - Departamento
Classificação. Subfilo Crustacea (número de espécies)
") Classificação Segundo Ruppert e Barnes, 1996 Subfilo Crustacea (número de espécies) Classe Remipedia (2) Classe Cephalocarida (9) Classe Branchiopoda (821) Classe Ostracoda (5650) Classe Copepoda (8405)
Classificação Segundo Ruppert e Barnes, 1996 Subfilo Crustacea (número de espécies) Classe Remipedia (2) Classe Cephalocarida (9) Classe Branchiopoda (821) Classe Ostracoda (5650) Classe Copepoda (8405)
Uma nova espécie de Phalloniscus Budde-Lund, 1908 (Crustacea, Oniscoidea) do sul do Brasil
 do sul do Brasil") REVISTA BRASILEIRA DE ZOOLOGIA Revta bras. Zool., 6 (4) : 6 1 1-6 1 5 31/XII/89 Uma nova espécie de Phalloniscus Budde-Lund, 1908 (Crustacea, Oniscoidea) do sul do Brasil ABSTRACT Cristina M. L. Zardo
REVISTA BRASILEIRA DE ZOOLOGIA Revta bras. Zool., 6 (4) : 6 1 1-6 1 5 31/XII/89 Uma nova espécie de Phalloniscus Budde-Lund, 1908 (Crustacea, Oniscoidea) do sul do Brasil ABSTRACT Cristina M. L. Zardo
Morfologia Externa Tórax e Abdome
 Morfologia Externa Tórax e Abdome Lucas Silva de Faria Morfologia Externa Revisão: Quais são os apêndices cefálicos??? Quais os tipos de aparelho bucal??? 1 Morfologia Externa Tórax 3 segmentos: Protórax
Morfologia Externa Tórax e Abdome Lucas Silva de Faria Morfologia Externa Revisão: Quais são os apêndices cefálicos??? Quais os tipos de aparelho bucal??? 1 Morfologia Externa Tórax 3 segmentos: Protórax
Papéis Avulsos de Zoologia
 HeMny, é- Hojn-yes; tf7 7 CRUSTACEA LIBRAR! IMITHSONIAN IHS*. RIIURN TO Vf-119 Papéis Avulsos de Zoologia PAPÉIS AVULSOS ZOOL., S. PAULO, 30 (19): 289-294 20.1.1977 SOBRE UMA NOVA ESPÉCIE BRASILEIRA DO
HeMny, é- Hojn-yes; tf7 7 CRUSTACEA LIBRAR! IMITHSONIAN IHS*. RIIURN TO Vf-119 Papéis Avulsos de Zoologia PAPÉIS AVULSOS ZOOL., S. PAULO, 30 (19): 289-294 20.1.1977 SOBRE UMA NOVA ESPÉCIE BRASILEIRA DO
A GNATHOSTOMATA DATA: 13/08/13 PROFS. ELEONORE SETZ, FELIPE TOLEDO
 BZ480 BZ681 ZOOLOGIA DE VERTEBRADOS AULA PRÁTICA: De CRANIATA A GNATHOSTOMATA DATA: 13/08/13 PROFS. ELEONORE SETZ, FELIPE TOLEDO e WESLEY SILVA COLABORAÇÃO: PAULO R. MANZANI MONITORES: Camila Castilho,
BZ480 BZ681 ZOOLOGIA DE VERTEBRADOS AULA PRÁTICA: De CRANIATA A GNATHOSTOMATA DATA: 13/08/13 PROFS. ELEONORE SETZ, FELIPE TOLEDO e WESLEY SILVA COLABORAÇÃO: PAULO R. MANZANI MONITORES: Camila Castilho,
Duas novas espécies de Mauesia (Cerambycidae, Lamiinae, Mauesini) e chave para identificação das espécies do gênero
 e chave para identificação das espécies do gênero") Volume 49(37):497 501, 2009 Duas novas espécies de Mauesia (Cerambycidae, Lamiinae, Mauesini) e chave para identificação das espécies do gênero Eleandro Moysés 1,2 Maria Helena M. Galileo 1,3 Abstract
Volume 49(37):497 501, 2009 Duas novas espécies de Mauesia (Cerambycidae, Lamiinae, Mauesini) e chave para identificação das espécies do gênero Eleandro Moysés 1,2 Maria Helena M. Galileo 1,3 Abstract
TÍTULO: ESTUDO AGONÍSTICO DE UMA NOVA ESPÉCIE DE PEIXE PIMELODELLA SP. NOV. POR MEIO DE FILMAGEM SUBAQUÁTICA.
 TÍTULO: ESTUDO AGONÍSTICO DE UMA NOVA ESPÉCIE DE PEIXE PIMELODELLA SP. NOV. POR MEIO DE FILMAGEM SUBAQUÁTICA. CATEGORIA: CONCLUÍDO ÁREA: CIÊNCIAS BIOLÓGICAS E SAÚDE SUBÁREA: CIÊNCIAS BIOLÓGICAS INSTITUIÇÃO:
TÍTULO: ESTUDO AGONÍSTICO DE UMA NOVA ESPÉCIE DE PEIXE PIMELODELLA SP. NOV. POR MEIO DE FILMAGEM SUBAQUÁTICA. CATEGORIA: CONCLUÍDO ÁREA: CIÊNCIAS BIOLÓGICAS E SAÚDE SUBÁREA: CIÊNCIAS BIOLÓGICAS INSTITUIÇÃO:
Ordem Gadiformes. Ana Cristina Teixeira Bonecker Claudia Akemi Pereira Namiki Márcia Salustiano de Castro Paula Nepomuceno Campos
 Ordem Gadiformes Ana Cristina Teixeira Bonecker Claudia Akemi Pereira Namiki Márcia Salustiano de Castro Paula Nepomuceno Campos SciELO Books / SciELO Livros / SciELO Libros BONECKER, ACT., et al. Ordem
Ordem Gadiformes Ana Cristina Teixeira Bonecker Claudia Akemi Pereira Namiki Márcia Salustiano de Castro Paula Nepomuceno Campos SciELO Books / SciELO Livros / SciELO Libros BONECKER, ACT., et al. Ordem
Crustáceos (crusta= crosta ou pele grossa)
") Grupo Mandibulata Subfilos: Miriápodes: quilópodes (centopéias e lacraias) e diplópodes (piolho de cobra). Crustáceos Insetos Novidades evolutivas: Mandíbulas (importantes para triturar alimentos); Olhos
Grupo Mandibulata Subfilos: Miriápodes: quilópodes (centopéias e lacraias) e diplópodes (piolho de cobra). Crustáceos Insetos Novidades evolutivas: Mandíbulas (importantes para triturar alimentos); Olhos
marsúpio (bolsa incubadora), localizado sob o tórax das fêmeas - é formado por grandes processos em forma de placas (oostegitos) das coxas torácicas.
, localizado sob o tórax das fêmeas - é formado por grandes processos em forma de placas (oostegitos) das coxas torácicas.") SUPERORDEM PERACARIDA CARACTERÍSTICAS MAIS DISTINTAS marsúpio (bolsa incubadora), localizado sob o tórax das fêmeas - é formado por grandes processos em forma de placas (oostegitos) das coxas torácicas.
SUPERORDEM PERACARIDA CARACTERÍSTICAS MAIS DISTINTAS marsúpio (bolsa incubadora), localizado sob o tórax das fêmeas - é formado por grandes processos em forma de placas (oostegitos) das coxas torácicas.
Os répteis, atualmente, abrangem cerca de 7 mil espécies conhecidas.
 Os répteis, atualmente, abrangem cerca de 7 mil espécies conhecidas. Eles surgiram há cerca de 300 milhões de anos, tendo provavelmente evoluído de certos anfíbios. Foram os primeiros vertebrados efetivamente
Os répteis, atualmente, abrangem cerca de 7 mil espécies conhecidas. Eles surgiram há cerca de 300 milhões de anos, tendo provavelmente evoluído de certos anfíbios. Foram os primeiros vertebrados efetivamente
RELATÓRIO DE MONITORAMENTO DA ICTIOFAUNA NA ÁREA DA UHE MAUÁ
 INSTITUTO DE TECNOLOGIA PARA O DESENVOLVIMENTO Centro de Hidráulica e Hidrologia Prof. Parigot de Souza RELATÓRIO DE MONITORAMENTO DA ICTIOFAUNA NA ÁREA DA UHE MAUÁ RELATÓRIO TÉCNICO Nº 39 2012 COORDENAÇÃO
INSTITUTO DE TECNOLOGIA PARA O DESENVOLVIMENTO Centro de Hidráulica e Hidrologia Prof. Parigot de Souza RELATÓRIO DE MONITORAMENTO DA ICTIOFAUNA NA ÁREA DA UHE MAUÁ RELATÓRIO TÉCNICO Nº 39 2012 COORDENAÇÃO
ACERCA DE UM SUB-HOLÓSTEO DO KARROO DE ANGOLA POR CARLOS TEIXEIRA
 ACERCA DE UM SUB-HOLÓSTEO DO KARROO DE ANGOLA POR CARLOS TEIXEIRA Em artigo anterior descrevi um curioso tipo de holósteo do Karroo de Angola (I}. No seguimento daquele estudo, ocupar-me-ei hoje da descrição
ACERCA DE UM SUB-HOLÓSTEO DO KARROO DE ANGOLA POR CARLOS TEIXEIRA Em artigo anterior descrevi um curioso tipo de holósteo do Karroo de Angola (I}. No seguimento daquele estudo, ocupar-me-ei hoje da descrição
GEOMORFOLOGIA FLUVIAL: PROCESSOS E FORMAS
 GEOMORFOLOGIA FLUVIAL: PROCESSOS E FORMAS Revista Brasileira de Geomorfologia Ano 9, número (2008) Salgado et al. Produção brasileira em revistas nacionais (200-2005) Produção brasileira em revistas internacionais
GEOMORFOLOGIA FLUVIAL: PROCESSOS E FORMAS Revista Brasileira de Geomorfologia Ano 9, número (2008) Salgado et al. Produção brasileira em revistas nacionais (200-2005) Produção brasileira em revistas internacionais
Registro de Phrynops williamsi no rio do Chapecó, Oeste de Santa Catarina, Brasil
 NOTA CIENTÍFICA Registro de Phrynops williamsi no rio do Chapecó, Oeste de Santa Catarina, Brasil SPIER, Edson Fernando * ; FAVRETTO, Mario Arthur ** ; ONGHERO JUNIOR, Osvaldo *** ; PIOVEZAN, Jean Carlos
NOTA CIENTÍFICA Registro de Phrynops williamsi no rio do Chapecó, Oeste de Santa Catarina, Brasil SPIER, Edson Fernando * ; FAVRETTO, Mario Arthur ** ; ONGHERO JUNIOR, Osvaldo *** ; PIOVEZAN, Jean Carlos
Ordem Beloniformes. Ana Cristina Teixeira Bonecker Claudia Akemi Pereira Namiki Márcia Salustiano de Castro Paula Nepomuceno Campos
 Ordem Beloniformes Ana Cristina Teixeira Bonecker Claudia Akemi Pereira Namiki Márcia Salustiano de Castro Paula Nepomuceno Campos SciELO Books / SciELO Livros / SciELO Libros BONECKER, ACT., et al. Ordem
Ordem Beloniformes Ana Cristina Teixeira Bonecker Claudia Akemi Pereira Namiki Márcia Salustiano de Castro Paula Nepomuceno Campos SciELO Books / SciELO Livros / SciELO Libros BONECKER, ACT., et al. Ordem
VINALTO GRAF ALICE FUMI KUMAGAI 3
 Acta Biol. Par., Curitiba, 33 (1, 2, 3, 4): 21-25. 2004 21 Uma espécie nova de Epirhyssa Cresson (Ichneumonidae, Hymenoptera) do Brasil 1 A new species of Epirhyssa Cresson, 1865 (Ichneumonidae, Hymenoptera)
Acta Biol. Par., Curitiba, 33 (1, 2, 3, 4): 21-25. 2004 21 Uma espécie nova de Epirhyssa Cresson (Ichneumonidae, Hymenoptera) do Brasil 1 A new species of Epirhyssa Cresson, 1865 (Ichneumonidae, Hymenoptera)
23) Schizodon fasciatus Spix & Agassiz, 1829
 Schizodon fasciatus Spix & Agassiz, 1829") 164 Ovos e larvas de peixes de água doce... 23) Schizodon fasciatus Spix & Agassiz, 1829 Foto: Dr. Hebert R. Axelrod - Fonte: Axelrod, 1991, p. 221. Nomes comuns: Boga, taguara, boga-lisa, araçu, aracu,
164 Ovos e larvas de peixes de água doce... 23) Schizodon fasciatus Spix & Agassiz, 1829 Foto: Dr. Hebert R. Axelrod - Fonte: Axelrod, 1991, p. 221. Nomes comuns: Boga, taguara, boga-lisa, araçu, aracu,
NOTA EXPLICATIVA DOS DADOS RECOLHIDOS NO ÂMBITO DOS TRABALHOS DE
 NOTA EXPLICATIVA DOS DADOS RECOLHIDOS NO ÂMBITO DOS TRABALHOS DE IMPLEMENTAÇÃO DA DIRECTIVA QUADRO DA ÁGUA A informação disponibilizada na página do Instituto da Água, I.P. (INAG) refere-se aos dados recolhidos
NOTA EXPLICATIVA DOS DADOS RECOLHIDOS NO ÂMBITO DOS TRABALHOS DE IMPLEMENTAÇÃO DA DIRECTIVA QUADRO DA ÁGUA A informação disponibilizada na página do Instituto da Água, I.P. (INAG) refere-se aos dados recolhidos
48) Pseudoplatystoma corruscans (Agassiz, 1829)
 Pseudoplatystoma corruscans (Agassiz, 1829)") 282 Ovos e larvas de peixes de água doce... 48) Pseudoplatystoma corruscans (Agassiz, 1829) Nomes comuns: Pintado, surubim, piracajara, pirá-pára, piracajiara e surubi. Distribuição geográfica: Bacias
282 Ovos e larvas de peixes de água doce... 48) Pseudoplatystoma corruscans (Agassiz, 1829) Nomes comuns: Pintado, surubim, piracajara, pirá-pára, piracajiara e surubi. Distribuição geográfica: Bacias
TAXONOMfA E DISTRIBUIÇÃO DO GÊNERO /,ucuer (DECAPODA, SERGESTOIDEA) NA COSTA CENTRAL BRASILEIRA
 NA COSTA CENTRAL BRASILEIRA") 1 RENE AZEVEDO CARDOSO TAXONOMfA E DISTRIBUIÇÃO DO GÊNERO /,ucuer (DECAPODA, SERGESTOIDEA) NA COSTA CENTRAL BRASILEIRA Dissertação apresentada à Coordenação de Pós-Graduação em Zoologia da Universidade
1 RENE AZEVEDO CARDOSO TAXONOMfA E DISTRIBUIÇÃO DO GÊNERO /,ucuer (DECAPODA, SERGESTOIDEA) NA COSTA CENTRAL BRASILEIRA Dissertação apresentada à Coordenação de Pós-Graduação em Zoologia da Universidade
60) Astronotus ocellatus (Cuvier, 1829)
 Astronotus ocellatus (Cuvier, 1829)") 336 Ovos e larvas de peixes de água doce... 60) Astronotus ocellatus (Cuvier, 1829) Nomes comuns : Acará-açu, apaiari e oscar. Ordem Perciformes - A. ocellatus 337 Distribuição geográfica: Originária da
336 Ovos e larvas de peixes de água doce... 60) Astronotus ocellatus (Cuvier, 1829) Nomes comuns : Acará-açu, apaiari e oscar. Ordem Perciformes - A. ocellatus 337 Distribuição geográfica: Originária da
Diversidade de Crustáceos
 Diversidade de Crustáceos Os primeiros crustáceos eram pequenos, marinhos que nadavam ou rastejam sobre o fundo Tronco com numerosos segmentos não fundidos com 1 par de apêndices birremes Classes: Remipedia
Diversidade de Crustáceos Os primeiros crustáceos eram pequenos, marinhos que nadavam ou rastejam sobre o fundo Tronco com numerosos segmentos não fundidos com 1 par de apêndices birremes Classes: Remipedia
04) Astyanax altiparanae Garutti & Britski, 2000
 Astyanax altiparanae Garutti & Britski, 2000") Ordem Characiformes - A. altiparanae 75 04) Astyanax altiparanae Garutti & Britski, 2000 Nomes comuns: Lambari, lambari-do-rabo-amarelo e tambiú. Distribuição geográfica: Bacia do alto rio Paraná (Garutti;
Ordem Characiformes - A. altiparanae 75 04) Astyanax altiparanae Garutti & Britski, 2000 Nomes comuns: Lambari, lambari-do-rabo-amarelo e tambiú. Distribuição geográfica: Bacia do alto rio Paraná (Garutti;
DIVISÃO DE GEOLOGIA E MINERALOGIA
 Número 65 MINISTÉRIO DA AGRICULTURA DEPARTAMENTO NACIONAL DA PRODUÇÃO MINERAL DIVISÃO DE GEOLOGIA E MINERALOGIA NOTAS PRELIMINARES E ESTUDOS RIO DE JANEIRO O GÊNERO PAULOCARIS CLARKE NAS CAMADAS II DO
Número 65 MINISTÉRIO DA AGRICULTURA DEPARTAMENTO NACIONAL DA PRODUÇÃO MINERAL DIVISÃO DE GEOLOGIA E MINERALOGIA NOTAS PRELIMINARES E ESTUDOS RIO DE JANEIRO O GÊNERO PAULOCARIS CLARKE NAS CAMADAS II DO
16) Piaractus mesopotamicus (Holmberg, 1887)
 Piaractus mesopotamicus (Holmberg, 1887)") 130 Ovos e larvas de peixes de água doce... 16) Piaractus mesopotamicus (Holmberg, 1887) Nomes comuns : Pacu e pacu-caranha. Distribuição geográfica: Bacia do rio da Prata (Romagosa; Paiva; Godinho; Storfer,
130 Ovos e larvas de peixes de água doce... 16) Piaractus mesopotamicus (Holmberg, 1887) Nomes comuns : Pacu e pacu-caranha. Distribuição geográfica: Bacia do rio da Prata (Romagosa; Paiva; Godinho; Storfer,
ALGUNS LANIATORES NOVOS DA REPUBLIC A ARGENTIN A MELLO-LEITÃ O
 r ALGUNS LANIATORES NOVOS DA REPUBLIC A ARGENTIN A MELLO-LEITÃ O Familia Falangodida s Subf amua, #,Trio ommatinas Genero :iiaoigalupo g. n. Comoro ocular muito elevado, com dois espinhos e situad o quasi
r ALGUNS LANIATORES NOVOS DA REPUBLIC A ARGENTIN A MELLO-LEITÃ O Familia Falangodida s Subf amua, #,Trio ommatinas Genero :iiaoigalupo g. n. Comoro ocular muito elevado, com dois espinhos e situad o quasi
CRUSTACEA. classes. Carapaça (dobra do tegumento; origem cefálica) Abdômen com menos de 9 segmentos
 Abdômen com menos de 9 segmentos") CRUSTACEA classes Malacostraca 5 6 1 3 2 5 4 Carapaça (dobra do tegumento; origem cefálica) Abdômen com menos de 9 segmentos 6 Maxillopoda Cephalocarida Remipedia Apêndices torácicos do tipo filopódio
CRUSTACEA classes Malacostraca 5 6 1 3 2 5 4 Carapaça (dobra do tegumento; origem cefálica) Abdômen com menos de 9 segmentos 6 Maxillopoda Cephalocarida Remipedia Apêndices torácicos do tipo filopódio
Características gerais
 Artrópodes Quem são? Muitas vezes, não percebemos a presença daqueles animais com corpos de formas estranhas e cores variadas, que vivem ao nosso redor, voam sobre nossas cabeças ou aqueles que se locomovem
Artrópodes Quem são? Muitas vezes, não percebemos a presença daqueles animais com corpos de formas estranhas e cores variadas, que vivem ao nosso redor, voam sobre nossas cabeças ou aqueles que se locomovem
UNIVERSIDADE FEDERAL DE ALAGOAS CAMPUS ARAPIRACA CIÊNCIAS BIOLÓGICAS - LICENCIATURA LUCIA VANESSA ROCHA SANTOS
 UNIVERSIDADE FEDERAL DE ALAGOAS CAMPUS ARAPIRACA CIÊNCIAS BIOLÓGICAS - LICENCIATURA LUCIA VANESSA ROCHA SANTOS DIVERSIDADE E DISTRIBUIÇÃO DOS CAMARÕES (CRUSTACEA PALAEMONIDAE E ATYIDAE) NO BAIXO RIO SÃO
UNIVERSIDADE FEDERAL DE ALAGOAS CAMPUS ARAPIRACA CIÊNCIAS BIOLÓGICAS - LICENCIATURA LUCIA VANESSA ROCHA SANTOS DIVERSIDADE E DISTRIBUIÇÃO DOS CAMARÕES (CRUSTACEA PALAEMONIDAE E ATYIDAE) NO BAIXO RIO SÃO
Professora Leonilda Brandão da Silva
 COLÉGIO ESTADUAL HELENA KOLODY E.M.P. TERRA BOA - PARANÁ Professora Leonilda Brandão da Silva E-mail: leonildabrandaosilva@gmail.com http://professoraleonilda.wordpress.com/ PROBLEMATIZAÇÃO Como você acha
COLÉGIO ESTADUAL HELENA KOLODY E.M.P. TERRA BOA - PARANÁ Professora Leonilda Brandão da Silva E-mail: leonildabrandaosilva@gmail.com http://professoraleonilda.wordpress.com/ PROBLEMATIZAÇÃO Como você acha
Lista de exercícios Aluno (a):
:") Lista de exercícios Aluno (a): Turma: 1ª série: Professor: Márcio França Antes de iniciar a lista de exercícios leia atentamente as seguintes orientações: É fundamental a apresentação de uma lista legível,
Lista de exercícios Aluno (a): Turma: 1ª série: Professor: Márcio França Antes de iniciar a lista de exercícios leia atentamente as seguintes orientações: É fundamental a apresentação de uma lista legível,
Revestimento corporal
 Répteis Répteis Os répteis (do latim reptare, 'rastejar') abrangem cerca de 7 mil espécies conhecidas. Eles surgiram há cerca de 300 milhões de anos, tendo provavelmente evoluído de certos anfíbios. Foram
Répteis Répteis Os répteis (do latim reptare, 'rastejar') abrangem cerca de 7 mil espécies conhecidas. Eles surgiram há cerca de 300 milhões de anos, tendo provavelmente evoluído de certos anfíbios. Foram
Filo Arthropoda Subfilo Crustacea
 Filo Arthropoda Subfilo Crustacea Etimologia: Latim: crusta = pele grossa ou crosta Importância: Ecológica: ocupam vários níveis tróficos - elos Algumas espécies são utilizadas em estudos ecotoxicológicos
Filo Arthropoda Subfilo Crustacea Etimologia: Latim: crusta = pele grossa ou crosta Importância: Ecológica: ocupam vários níveis tróficos - elos Algumas espécies são utilizadas em estudos ecotoxicológicos
Acta Biol. Par., Curitiba, 32 (1, 2, 3, 4):
:") Acta Biol. Par., Curitiba, 32 (1, 2, 3, 4): 83-94. 2003 83 Lanocira kroyeri Hansen (Isopoda: Corallanidae) redescrição e nova ocorrência para a Ilha Arvoredo, Santa Catarina, Brasil ¹ Lanocira kroyeri
Acta Biol. Par., Curitiba, 32 (1, 2, 3, 4): 83-94. 2003 83 Lanocira kroyeri Hansen (Isopoda: Corallanidae) redescrição e nova ocorrência para a Ilha Arvoredo, Santa Catarina, Brasil ¹ Lanocira kroyeri
Paguroidea, eles formam, junto com o própodo, estruturas subqueladas ou queladas, com escamas córneas "raspadoras" na face ventral.
 Paguroidea, eles formam, junto com o própodo, estruturas subqueladas ou queladas, com escamas córneas "raspadoras" na face ventral. Os Thalassinidea formam um grupo de famílias, gêneros e espécies, cujas
Paguroidea, eles formam, junto com o própodo, estruturas subqueladas ou queladas, com escamas córneas "raspadoras" na face ventral. Os Thalassinidea formam um grupo de famílias, gêneros e espécies, cujas
OCORRÊNCIA DOS CAMARÕES Macrobrachium amazonicum (HELLER, 1862) E Macrobrachium jelskii (MIERS, 1877) Resumo
 E Macrobrachium jelskii (MIERS, 1877) Resumo") Universidade Regional do Cariri URCA Caderno de Cultura e Ciência, Ano XI, v.15, n.1, Out, 2016 ISSN 1980-5861 OCORRÊNCIA DOS CAMARÕES Macrobrachium amazonicum (HELLER, 1862) E Macrobrachium jelskii (MIERS,
Universidade Regional do Cariri URCA Caderno de Cultura e Ciência, Ano XI, v.15, n.1, Out, 2016 ISSN 1980-5861 OCORRÊNCIA DOS CAMARÕES Macrobrachium amazonicum (HELLER, 1862) E Macrobrachium jelskii (MIERS,
ZOOLOGIA DE VERTEBRADOS
 ZOOLOGIA DE VERTEBRADOS CURSO: Ciências Biológicas 3º Ano 2º semestre Campo 2 - Anfíbios Preparação para o campo Professores: Virgínia Sanches Uieda e Wilson Uieda Colaborador: Silvio César de Almeida
ZOOLOGIA DE VERTEBRADOS CURSO: Ciências Biológicas 3º Ano 2º semestre Campo 2 - Anfíbios Preparação para o campo Professores: Virgínia Sanches Uieda e Wilson Uieda Colaborador: Silvio César de Almeida
BIODIVERSIDADE DE MACROINVERTEBRADOS BENTÔNICOS DO LITORAL DO ESTADO DA BAHIA
 TÍTULO DO RESUMO BIODIVERSIDADE DE MACROINVERTEBRADOS BENTÔNICOS DO LITORAL DO ESTADO DA BAHIA 1. Bolsista FAPESB/CNPq, Verônica Santos de Andrade Graduanda em Ciências Biológicas, Universidade Estadual
TÍTULO DO RESUMO BIODIVERSIDADE DE MACROINVERTEBRADOS BENTÔNICOS DO LITORAL DO ESTADO DA BAHIA 1. Bolsista FAPESB/CNPq, Verônica Santos de Andrade Graduanda em Ciências Biológicas, Universidade Estadual
ESTUDO DA OCORRÊNCIA DA ICTIOFAUNA EM SANTA CATARINA
 ESTUDO DA OCORRÊNCIA DA ICTIOFAUNA EM SANTA CATARINA Modalidade: ( ) Ensino ( x ) Pesquisa ( ) Extensão Nível: ( x ) Médio ( ) Superior ( ) Pós-graduação Área: ( ) Química ( ) Informática ( ) Ciências
ESTUDO DA OCORRÊNCIA DA ICTIOFAUNA EM SANTA CATARINA Modalidade: ( ) Ensino ( x ) Pesquisa ( ) Extensão Nível: ( x ) Médio ( ) Superior ( ) Pós-graduação Área: ( ) Química ( ) Informática ( ) Ciências
ALIMENTAÇÃO DE Rachycentron canadum NA BAÍA DE TODOS OS SANTOS
 ALIMENTAÇÃO DE Rachycentron canadum NA BAÍA DE TODOS OS SANTOS Lopes, P.R.D. (1) ; Oliveira-Silva, J.T. (1) ; Sena, M.P. (1) andarilho40@gmail.com (1) Departamento de Ciências Biológicas da Universidade
ALIMENTAÇÃO DE Rachycentron canadum NA BAÍA DE TODOS OS SANTOS Lopes, P.R.D. (1) ; Oliveira-Silva, J.T. (1) ; Sena, M.P. (1) andarilho40@gmail.com (1) Departamento de Ciências Biológicas da Universidade
Ordem Tetraodontiformes
 Ordem Tetraodontiformes Ana Cristina Teixeira Bonecker Claudia Akemi Pereira Namiki Márcia Salustiano de Castro Paula Nepomuceno Campos SciELO Books / SciELO Livros / SciELO Libros BONECKER, ACT., et al.
Ordem Tetraodontiformes Ana Cristina Teixeira Bonecker Claudia Akemi Pereira Namiki Márcia Salustiano de Castro Paula Nepomuceno Campos SciELO Books / SciELO Livros / SciELO Libros BONECKER, ACT., et al.
Terrário miniatura do Planeta Terra 1
 A TEORIA EM PRÁTICA. EXEMPLO DE ATIVIDADE Terrário miniatura do Planeta Terra 1 A montagem de um terrário é uma tarefa simples e um rico instrumento pedagógico. Sua confecção permite trabalhar o reaproveitamento
A TEORIA EM PRÁTICA. EXEMPLO DE ATIVIDADE Terrário miniatura do Planeta Terra 1 A montagem de um terrário é uma tarefa simples e um rico instrumento pedagógico. Sua confecção permite trabalhar o reaproveitamento
CARACTERÍSTICAS DO MEIO AQUÁTICO
 ÁGUAS CONTINENTAIS CARACTERÍSTICAS DO MEIO AQUÁTICO # Alta capacidade para solubilização de compostos orgânicos e inorgânicos. # Gradientes verticais e, em certos casos, gradientes horizontais, que se
ÁGUAS CONTINENTAIS CARACTERÍSTICAS DO MEIO AQUÁTICO # Alta capacidade para solubilização de compostos orgânicos e inorgânicos. # Gradientes verticais e, em certos casos, gradientes horizontais, que se
DEPARTAMENTO DE ZOOLOGIA. DUAS NOVAS ESPÉCIES DiE OPILIÕES DO ESiTANDO DO ESPíRITO SANTO (*) POR INTRODUÇÁO
 POR INTRODUÇÁO") Vol. V, N." 30 - pp. 281-286 18-VI-1945 PAPÉI s AVULSOS DO DEPARTAMENTO DE ZOOLOGIA SECRETARIA DA AGRICULTURA - S. PAULO - BRASIL DUAS NOVAS ESPÉCIES DiE OPILIÕES DO ESiTANDO DO ESPíRITO SANTO (*) POR
Vol. V, N." 30 - pp. 281-286 18-VI-1945 PAPÉI s AVULSOS DO DEPARTAMENTO DE ZOOLOGIA SECRETARIA DA AGRICULTURA - S. PAULO - BRASIL DUAS NOVAS ESPÉCIES DiE OPILIÕES DO ESiTANDO DO ESPíRITO SANTO (*) POR
= H. azteca (Saussure)]. Hyalella curvispina Shoemaker, 1942: 79 - Barnard, 1958: 79; Cavalieri, 1968: 116; Holsinger, 1981: 39.
![= H. azteca (Saussure)]. Hyalella curvispina Shoemaker, 1942: 79 - Barnard, 1958: 79; Cavalieri, 1968: 116; Holsinger, 1981: 39.](/thumbs/53/32300308.jpg "= H. azteca (Saussure)]. Hyalella curvispina Shoemaker, 1942: 79 - Barnard, 1958: 79; Cavalieri, 1968: 116; Holsinger, 1981: 39.") REVISTA BRASILEIRA DE ZOOLOGIA Revta bras. Zool., S Paulo 3(4): 209 217 28.vi.1985 REDESCRIÇÃO DE HY ALELLA PERNIX (MOREIRA) (AMPHIPODA - HYALELLIDAE) COM DISCUSSÃO DE SEU SINONIMO H. CURVISPINA SHOEMAKER
REVISTA BRASILEIRA DE ZOOLOGIA Revta bras. Zool., S Paulo 3(4): 209 217 28.vi.1985 REDESCRIÇÃO DE HY ALELLA PERNIX (MOREIRA) (AMPHIPODA - HYALELLIDAE) COM DISCUSSÃO DE SEU SINONIMO H. CURVISPINA SHOEMAKER
BIOMAS. Professora Débora Lia Ciências/Biologia
 BIOMAS Professora Débora Lia Ciências/Biologia BIOMAS - Aquáticos Mares e oceanos Talássicos São as regiões com a maior variedade de vida do planeta, nem as florestas tropicais igualam-se às regiões litorâneas;
BIOMAS Professora Débora Lia Ciências/Biologia BIOMAS - Aquáticos Mares e oceanos Talássicos São as regiões com a maior variedade de vida do planeta, nem as florestas tropicais igualam-se às regiões litorâneas;
Biodiversidade marinha e lacustre
 Biodiversidade marinha e lacustre Lagostim Tenho duas pinças, antenas e sofro mudanças de carapaça. Para me proteger, fico totalmente escondido. Na escolha do meu habitat, tenho em atenção o tipo de substrato
Biodiversidade marinha e lacustre Lagostim Tenho duas pinças, antenas e sofro mudanças de carapaça. Para me proteger, fico totalmente escondido. Na escolha do meu habitat, tenho em atenção o tipo de substrato
CONFEDERAÇÃO BRASILEIRA DE CINOFILIA Fédération Cynologique Internationale GRUPO 7. Padrão FCI 1 07/09/1998
 CONFEDERAÇÃO BRASILEIRA DE CINOFILIA Fédération Cynologique Internationale GRUPO 7 Padrão FCI 1 07/09/1998 Padrão Oficial da Raça POINTER INGLÊS ENGLISH POINTER CONFEDERAÇÃO BRASILEIRA DE CINOFILIA Classificação
CONFEDERAÇÃO BRASILEIRA DE CINOFILIA Fédération Cynologique Internationale GRUPO 7 Padrão FCI 1 07/09/1998 Padrão Oficial da Raça POINTER INGLÊS ENGLISH POINTER CONFEDERAÇÃO BRASILEIRA DE CINOFILIA Classificação
Espécies novas de Eucerinae Neotropicais e notas sobre Dasyhalonia phaeoptera Moure & Michener (Hymenoptera, Anthophoridae) 1
 1") Espécies novas de Eucerinae Neotropicais e notas sobre Dasyhalonia phaeoptera Moure & Michener (Hymenoptera, Anthophoridae) 1 Danúncia Urban 2 ABSTRACT. New species of neotropical Eucerinae and notes on
Espécies novas de Eucerinae Neotropicais e notas sobre Dasyhalonia phaeoptera Moure & Michener (Hymenoptera, Anthophoridae) 1 Danúncia Urban 2 ABSTRACT. New species of neotropical Eucerinae and notes on
Anexo ao documento orientador do MS Fichas Técnicas explicativas das provas.
 ANEXOS Anexo ao documento orientador do MS 2018 1 Tabelas de referência; 2 - Fichas Técnicas explicativas das provas. 2 Tabela de Referência Mega Sprinter (40 metros) FEMININO MASCULINO INFANTIS A INFANTIS
ANEXOS Anexo ao documento orientador do MS 2018 1 Tabelas de referência; 2 - Fichas Técnicas explicativas das provas. 2 Tabela de Referência Mega Sprinter (40 metros) FEMININO MASCULINO INFANTIS A INFANTIS
F. Chave ilustrada para ordens
 MILLI-PEET; Illustrated Key to Order; Page - 1 - F. Chave ilustrada para ordens 1A Parede do corpo mole; tergitos com tufos de cerdas plumosas, com um par de tufos formados por cerdas longas (Fig. 8) na
MILLI-PEET; Illustrated Key to Order; Page - 1 - F. Chave ilustrada para ordens 1A Parede do corpo mole; tergitos com tufos de cerdas plumosas, com um par de tufos formados por cerdas longas (Fig. 8) na
Livro Vermelho dos Crustáceos do Brasil: Avaliação ISBN SBC AVALIAÇÃO DOS CAMARÕES ATIÍDEOS (DECAPODA: ATYIDAE)
") Livro Vermelho dos Crustáceos do Brasil: Avaliação 2010-2014 ISBN 978-85-93003-00-4 2016 SBC CAPÍTULO 5 AVALIAÇÃO DOS CAMARÕES ATIÍDEOS (DECAPODA: ATYIDAE) Fernando L. Mantelatto, Lucas S. Torati, Leonardo
Livro Vermelho dos Crustáceos do Brasil: Avaliação 2010-2014 ISBN 978-85-93003-00-4 2016 SBC CAPÍTULO 5 AVALIAÇÃO DOS CAMARÕES ATIÍDEOS (DECAPODA: ATYIDAE) Fernando L. Mantelatto, Lucas S. Torati, Leonardo
Filo Arthropoda Subfilo Crustacea
 Filo Arthropoda Subfilo Crustacea Etimologia: Latim: crusta pele grossa ou crosta Importância: Ecológica: ocupam vários níveis tróficos Algumas espécies são utilizadas em estudos ecotoxicológicos Indicadores
Filo Arthropoda Subfilo Crustacea Etimologia: Latim: crusta pele grossa ou crosta Importância: Ecológica: ocupam vários níveis tróficos Algumas espécies são utilizadas em estudos ecotoxicológicos Indicadores
PAPEIS AVULSOS DEPARTAMENTO DE ZOOLOGIA SECRETARIA DA AGRICULTURA- S. PAULO - BRASIL POR
 Vol,. V, N. 0 28 - pp. 271-276 18-VI-1945 PAPEIS AVULSOS DO DEPARTAMENTO DE ZOOLOGIA SECRETARIA DA AGRICULTURA- S. PAULO - BRASIL OPILlõES DA CHÃCAHA "DR. L. J. LANE", EM SÃO PAULO (CAPITAL) (*) POR B.
Vol,. V, N. 0 28 - pp. 271-276 18-VI-1945 PAPEIS AVULSOS DO DEPARTAMENTO DE ZOOLOGIA SECRETARIA DA AGRICULTURA- S. PAULO - BRASIL OPILlõES DA CHÃCAHA "DR. L. J. LANE", EM SÃO PAULO (CAPITAL) (*) POR B.
Lagostas. Cavacos. J Fontes ImagDOP. F Cardigos - ImagDOP
 Lagostas J Fontes ImagDOP As lagostas (Palinurus elephas) podem ser encontradas entre a superfície e os 70 metros de profundidade. Os sexos estão separados (ou seja, existem indivíduos machos e fêmeas)
Lagostas J Fontes ImagDOP As lagostas (Palinurus elephas) podem ser encontradas entre a superfície e os 70 metros de profundidade. Os sexos estão separados (ou seja, existem indivíduos machos e fêmeas)
RELATÓRIO DE MONITORAMENTO DA ICTIOFAUNA NA ÁREA DA UHE MAUÁ
 INSTITUTO DE TECNOLOGIA PARA O DESENVOLVIMENTO Centro de Hidráulica e Hidrologia Prof. Parigot de Souza RELATÓRIO DE MONITORAMENTO DA ICTIOFAUNA NA ÁREA DA UHE MAUÁ RELATÓRIO TÉCNICO Nº 39 2012 COORDENAÇÃO
INSTITUTO DE TECNOLOGIA PARA O DESENVOLVIMENTO Centro de Hidráulica e Hidrologia Prof. Parigot de Souza RELATÓRIO DE MONITORAMENTO DA ICTIOFAUNA NA ÁREA DA UHE MAUÁ RELATÓRIO TÉCNICO Nº 39 2012 COORDENAÇÃO
Arthropoda: Características gerais Crustacea Myriapoda. Marina Ferraz de Camargo Barbosa. LEA0498 Piracicaba, 08 de Agosto de 2018
 Arthropoda: Características gerais Crustacea Myriapoda Marina Ferraz de Camargo Barbosa LEA0498 Piracicaba, 08 de Agosto de 2018 Classificação dos Artrópodes (de acordo com Brusca et al., 2018) Caracterização
Arthropoda: Características gerais Crustacea Myriapoda Marina Ferraz de Camargo Barbosa LEA0498 Piracicaba, 08 de Agosto de 2018 Classificação dos Artrópodes (de acordo com Brusca et al., 2018) Caracterização
A profundidade do oceano é de 3794 m (em média), mais de cinco vezes a altura média dos continentes.
, mais de cinco vezes a altura média dos continentes.") Hidrosfera Compreende todos os rios, lagos,lagoas e mares e todas as águas subterrâneas, bem como as águas marinhas e salobras, águas glaciais e lençóis de gelo, vapor de água, as quais correspondem a
Hidrosfera Compreende todos os rios, lagos,lagoas e mares e todas as águas subterrâneas, bem como as águas marinhas e salobras, águas glaciais e lençóis de gelo, vapor de água, as quais correspondem a
Orogênese (formação de montanhas): o choque entre placas tectônicas forma as cordilheiras.
: o choque entre placas tectônicas forma as cordilheiras.") O RELEVO TERRESTRE A superfície terrestre não é uniforme, mas sim composta por irregularidades, apresentando formas altas ou baixas, planas ou onduladas. O relevo pode ser definido como o conjunto das
O RELEVO TERRESTRE A superfície terrestre não é uniforme, mas sim composta por irregularidades, apresentando formas altas ou baixas, planas ou onduladas. O relevo pode ser definido como o conjunto das
A Bacia Amazônica: Conectividade, Migrações e Ciência Cidadã Conhecendo os peixes migradores
 A Bacia Amazônica: Conectividade, Migrações e Ciência Cidadã Conhecendo os peixes migradores Esta apresentação foi organizada a partir de informações disponíveis em http://pt.aguasamazonicas.org/fish/
A Bacia Amazônica: Conectividade, Migrações e Ciência Cidadã Conhecendo os peixes migradores Esta apresentação foi organizada a partir de informações disponíveis em http://pt.aguasamazonicas.org/fish/
Parte IV - Répteis e Anfíbios / Reptiles and Amphibians 1932 uma nova espécie de Cyclorhamphus
 Parte IV - Répteis e Anfíbios / Reptiles and Amphibians 1932 uma nova espécie de Cyclorhamphus Adolpho Lutz SciELO Books / SciELO Livros / SciELO Libros BENCHIMOL, JL., and SÁ, MR., eds. and orgs. Adolpho
Parte IV - Répteis e Anfíbios / Reptiles and Amphibians 1932 uma nova espécie de Cyclorhamphus Adolpho Lutz SciELO Books / SciELO Livros / SciELO Libros BENCHIMOL, JL., and SÁ, MR., eds. and orgs. Adolpho
Joaquim Olinto Branco* & Maria Jose Lunardon-Branco** ABSTRACT INTRODUÇÃO
 CRESCIMENTO E TAMANHO DE PRIMEIRA MATURAÇÃO EM Callinectes ornatus ORDWAY, 1863 (DECAPODA, PORTUNIDAE) DA REGIÃO DE MATINHOS, PARANÁ, BRASIL. Joaquim Olinto Branco* & Maria Jose Lunardon-Branco** Núcleo
CRESCIMENTO E TAMANHO DE PRIMEIRA MATURAÇÃO EM Callinectes ornatus ORDWAY, 1863 (DECAPODA, PORTUNIDAE) DA REGIÃO DE MATINHOS, PARANÁ, BRASIL. Joaquim Olinto Branco* & Maria Jose Lunardon-Branco** Núcleo
QUARTA A OITAVA SEMANA DO DESENVOLVIMENTO EMBRIONÁRIO
 QUARTA A OITAVA SEMANA DO DESENVOLVIMENTO EMBRIONÁRIO ASPECTOS INTERNOS E EXTERNOS DO EMBRIÃO 1 O PERÍODO DE ORGANOGÊNESE OCORRE DA QUARTA À OITAVA SEMANA DO DESENVOLVIMENTO EMBRIONÁRIO. AO FINAL DA OITAVA
QUARTA A OITAVA SEMANA DO DESENVOLVIMENTO EMBRIONÁRIO ASPECTOS INTERNOS E EXTERNOS DO EMBRIÃO 1 O PERÍODO DE ORGANOGÊNESE OCORRE DA QUARTA À OITAVA SEMANA DO DESENVOLVIMENTO EMBRIONÁRIO. AO FINAL DA OITAVA
A utilização do método de flutuadores na medição da vazão do rio Biguaçu no Estado de Santa Catarina
 A utilização do método de flutuadores na medição da vazão do rio Biguaçu no Estado de Santa Catarina Vicente Rocha Silva Centro Universitário Municipal de São José (SC) USJ E-mail: vicenters@usp.br RESUMO
A utilização do método de flutuadores na medição da vazão do rio Biguaçu no Estado de Santa Catarina Vicente Rocha Silva Centro Universitário Municipal de São José (SC) USJ E-mail: vicenters@usp.br RESUMO
Ecologia populacional e reprodutiva de Potimirim glabra (Kingsley, 1954) (Caridea, Atyidae) em riachos da região de Ubatuba-SP.
 (Caridea, Atyidae) em riachos da região de Ubatuba-SP.") Ecologia populacional e reprodutiva de Potimirim glabra (Kingsley, 1954) (Caridea, Atyidae) em riachos da região de Ubatuba-SP. Patrícia Hoffmann Orientadora: Profa. Dra. Maria Lucia Negreiros Fransozo
Ecologia populacional e reprodutiva de Potimirim glabra (Kingsley, 1954) (Caridea, Atyidae) em riachos da região de Ubatuba-SP. Patrícia Hoffmann Orientadora: Profa. Dra. Maria Lucia Negreiros Fransozo
2.1 ECOSSISTEMAS RIBEIRINHOS - O RIO
 2.1 ECOSSISTEMAS RIBEIRINHOS - O RIO FICHA DE CAMPO 1 1/6 FICHA DE REGISTO DA CARACTERIZAÇÃO DO TROÇO DE UM RIO Local Data Nome Localização (Google Earth) Latitude º Longitude º Curso de água Referência
2.1 ECOSSISTEMAS RIBEIRINHOS - O RIO FICHA DE CAMPO 1 1/6 FICHA DE REGISTO DA CARACTERIZAÇÃO DO TROÇO DE UM RIO Local Data Nome Localização (Google Earth) Latitude º Longitude º Curso de água Referência