HISTÓRIA NATURAL E MORFOLOGIA DE Heraclides anchisiades capys (HÜBNER, [1809]) (LEPIDOPTERA : PAPILIONIDAE)
|
|
|
- Lucinda Cipriano
- 5 Há anos
- Visualizações:
Transcrição
1 LUIS ANDERSON RIBEIRO LEITE HISTÓRIA NATURAL E MORFOLOGIA DE Heraclides anchisiades capys (HÜBNER, [1809]) (LEPIDOPTERA : PAPILIONIDAE) Dissertação apresentada à Coordenação do Curso de Pós-Graduação em Ciências Biológicas, Área de Concentração em Entomologia, da Universidade Federal do Paraná, para a obtenção do título de Mestre em Ciências Biológicas. CURITIBA Julho/2009
2 Livros Grátis Milhares de livros grátis para download.
3 II
4 Dedico, aos meus pais e irmãs pelo amor incondicional e apoio em todos os momentos até então, mas especialmente à Ludmilla Meyer Montenegro, por todo carinho, companheirismo e paciência durante todos estes anos de convivência. III
5 AGRADECIMENTOS À Profª. Drª. Mirna Martins Casagrande, pela orientação, amizade e pela confiança depositada em mim neste período de minha vida e ao Prof. Dr. Olaf Hermann Hendrik Mielke, pela co-orientação e pelos valiosos ensinamentos provindos de seus muitos anos de experiência. Ao Programa de Pós-graduação em Entomologia do Departamento de Zoologia da Universidade Federal do Paraná (UFPR) pela oportunidade; ao CNPq pela concessão da bolsa, sem a qual este trabalho não seria possível; ao Centro de Microscopia Eletrônica e ao Táxon line da Universidade Federal do Paraná, ambos pela elaboração de fotografias. À Profª. Drª. Danuncia Urban, pela amizade cultivada nestes dois anos e por se mostrar sempre atenciosa e disposta a solucionar as dúvidas surgidas diariamente. À Profª Drª Angélica Maria Penteado Dias da Universidade Federal de São Carlos pela disponibilidade e atenção quanto à identificação de material. Aos companheiros do Laboratório de Estudos de Lepidoptera Neotropical, em especial a Gustavo Cavalheiro Amarília e Marina Moraes Carlos pela amizade acima de tudo, pelas trocas de informações diárias e pelos auxílios nas horas mais importantes. Aos professores e amigos de curso do programa de Pós-Graduação em Entomologia da Universidade Federal do Paraná, pelos ensinamentos e pela boa convivência em todo este tempo. esperanças. Aos amigos e familiares que me apoiam e depositam em mim grandes IV
6 The scientist does not study nature because it is useful to do so. He studies it because he takes pleasure in it; and he takes pleasure in it because it is beautiful. If nature were not beautiful, it would not be worth knowing and life would not be worth living. (Henri Poincaré). V
7 SUMÁRIO Resumo Geral Abstract Capítulo I Resumo Abstract Introdução Objetivo Material e Métodos Resultados Cabeça Apêndices cefálicos Região Cervical Tórax Apêndices torácicos Abdome Discussão Figuras Referências Bibliográficas Capítulo II Resumo Abstract Introdução Objetivos Material e Métodos Resultados Aspectos Morfológicos Ovo Larva 1º ínstar Larva 2º ínstar Larva 3º ínstar Larva 4º ínstar Larva 5º ínstar Larva 6º ínstar Pupa Comportamento, mecanismos de defesa e parasitismo Discussão Figuras Referências Bibliográficas VI
8 LISTA DE FIGURAS CAPÍTULO I Figura 1 - Macho em vista dorsal Figura 2 - Macho em vista ventral Figura 3 - Fêmea em vista dorsal Figura 4 - Fêmea em vista ventral Figura 5 - Cabeça - vista frontal Figura 6 - Cabeça - vista ventral Figura 7 - Detalhe da vista ventral da cabeça Figura 8 - Cabeça - vista posterior Figura 9 - Cabeça - vista dorsal Figura 10 - Cabeça vista lateral Figura 11 - Antena Figura 12 - Palpo labial vista externa Figura 13 - Palpo labial vista interna Figura 14 - Região cervical vista lateral Figura 15 - Região cervical vista ventral Figura 16 - Escapo Figura 17 - Detalhe da área sensitiva do escapo Figura 18 - Artículo distal da antena Figura 19 - Aparelho bucal vista ventral Figura 20 - Artículo distal do palpo Figura 21 - Cardo Figura 22 - Palpo maxilar rudimentar Figura 23 - Ápice da gálea Figura 24 - Ápice da gálea Figura 25 - Ápice da gálea Figura 26 - Região do pilífero e rudimento mandibular Figura 27 - Região do pilífero e rudimento mandibular Figura 28 - Fóvea tentorial anterior Figura 29 - Fóvea tentorial posterior Figura 30 - Quetosema Figura 31 - Tórax - vista dorsal Figura 32 - Protórax - vista dorsal Figura 33 - Tórax - vista lateral Figura 34 - Protórax - vista ventral Figura 35 - Mesotórax e Metatórax - vista ventral Figura 36 - Escleritos axilares - vista dorsal Figura 37 - Perna protorácica Figura 38 - Epífise Figura 39 - Detalhe do último tarsômero protorácico Figura 40 - Perna mesotorácica Figura 41 - Esporão tibial Figura 42 - Perna metatorácica Figura 43 - Tégula - vista lateral anterior Figura 44 - Tégula - vista lateral posterior Figura 45 - Asas do macho Figura 46 - Asas da fêmea VII
9 Figura 47 - Trocantim e extremidade basal da coxa I Figura 48 - Trocantim e extremidade basal da coxa I Figura 49 - Articulação coxa-pleural Figura 50 - Epífise Figura 51 - Epífise Figura 52 - Distitarso - vista lateral Figura 53 - Distitarso - vista lateral Figura 54 - Distitarso - vista ventral Figura 55 - Distitarso - vista ventral Figura 56 - Basisterno I Figura 57 - Almofada do basalar III Figura 58 - Tégula vista geral Figura 59 - Tégula vista da região de cerdas Figura 60 - Abdome do macho - vista lateral Figura 61 - Abdome do macho - vista ventral Figura 62 - Abdome da fêmea - vista lateral Figura 63 - Abdome da fêmea - vista ventral Figura 64 - Genitália masculina - vista lateral Figura 65 - Genitália masculina - vista posterior Figura 66 - Valva Figura 67 - Edeago - vista lateral anterior Figura 68 - Edeago - vista lateral posterior Figura 69 - Genitália feminina - vista ventral Figura 70 - Genitália feminina em perfil VIII
10 LISTA DE FIGURAS CAPÍTULO II Figura 1 - Ovo - Imagem da postura Figura 2 - Ovo - Imagem da postura Figura 3 - Ovo - vista lateral Figura 4 - Ovo - vista lateral Figura 5 - Ovo - vista lateral Figura 6 - Ovo - vista superior Figura 7 - Ovo - vista superior Figura 8 - Larva de 1º ínstar - vista lateral Figura 9 - Larva de 1º ínstar cápsula cefálica - vista frontal Figura 10 - Larva de 1º ínstar cápsula cefálica - vista frontal Figura 11 - Cápsula cefálica 1º ínstar Figura 12 - Larva de 1º ínstar - quetotaxia do tórax Figura 13 - Larva de 1º ínstar - quetotaxia do abdome Figura 14 - Larva de 1º ínstar Figura 15 - Larva de 1º ínstar Figura 16 - Larva de 2º ínstar Figura 17 - Larva de 2º ínstar Figura 18 - Larva de 3º ínstar Figura 19 - Larva de 3º ínstar Figura 20 - Larva de 4º ínstar Figura 21 - Larva de 4º ínstar Figura 22 - Larva de 5º ínstar Figura 23 - Larva de 5º ínstar Figura 24 - Larva de 6º ínstar Figura 25 - Larva de 6º ínstar Figura 26 - Pré-pupa na planta hospedeira Figura 27 - Pré-pupa em concreto Figura 28 - Pupa - padrão esverdeado Figura 29 - Pupa - padrão castanho-escuro Figura 30 - Pupa - padrão pardo-acinzentado Figura 31 - Pupa Figura 32 - Pupa Figura 33 - Pupa Figura 34 - Pupa Figura 35 - Pupa Figura 36 - Pupa Figura 37 - Pupa Figura 38 - Pupa Figura 39 - Larva de 5º ínstar - estemas Figura 40 - Larva de 5º ínstar - detalhe de um estema Figura 41 - Larva de 6º ínstar - cápsula cefálica vista ventral Figura 42 - Larva de 6º ínstar - região inferior da cápsula cefálica Figura 43 - Fiandeira Figura 44 - Antena Figura 45 - Larva de 6º ínstar - perna protorácica Figura 46 - Larva de 6º ínstar - perna protorácica IX
11 Figura 47 - Larva de 6º ínstar - perna abdominal A Figura 48 - Larva de 6º ínstar - perna abdominal A3 - A Figura 49 - Larva de 6º ínstar - perna abdominal A3 - A Figura 50 - Larva de 6º ínstar - gancho da perna abdominal Figura 51 - Larva de 6º ínstar - quetotaxia tórax Figura 52 - Larva de 6º ínstar - quetotaxia abdome Figura 53 - Larva de 6º ínstar - cabeça Figura 54 - Larva de 6º ínstar - osmetério Figura 55 - Larva de 6º ínstar - osmetério Figura 56 - Larva de 6º ínstar - osmetério Figura 57 - Larva de 6º ínstar - osmetério Figura 58 - Larva de 6º ínstar - osmetério Figura 59 - Larva de 6º ínstar - espiráculo abdominal Figura 60 - Pupa - superfície ventral da cabeça Figura 61 - Pupa - olho Figura 62 - Pupa - espiráculo abdominal Figura 63 - Pupa - porção distal do abdome Figura 64 - Pupa - ganchos do cremaster Figura 65 - Pupa - ganchos do cremaster Figura 66 - Pupa - macho esternos VII a X Figura 67 - Pupa - fêmea esternos VII a X Figura 68 - Pupa - vista dorsal Figura 69 - Pupa - vista ventral Figura 70 - Pupa - vista lateral Figura 71 - Exúvia pupal e Pedinopelte gravenstii Figura 72 - Exúvia pupal e Pedinopelte gravenstii X
12 HISTÓRIA NATURAL E MORFOLOGIA DE Heraclides anchisiades capys (Hübner, [1809]) (LEPIDOPTERA : PAPILIONIDAE) RESUMO Papilionidae inclui cerca de 600 espécies, as quais são predominantemente borboletas tropicais. Uma subespécie muito comum dentro dessa família é Heraclides anchisiades capys, borboleta de coloração escura que se alimenta de Citrus spp. (Rutaceae). Possui grande potencial econômico e facilidade de criação em cativeiro, contudo, até os dias atuais são raras as informações sobre sua biologia. O propósito deste estudo é evidenciar as estruturas morfológicas dos adultos e imaturos da espécie, além dos aspectos relacionados à história natural, visando um enfoque comparativo com os demais Papilionoidea e Hesperioidea estudados. As estruturas dos adultos foram removidas do corpo, entretanto, seus apêndices isolados e fervidos anteriormente em hidróxido de potássio (KOH) 10% para clarificar e facilitar a remoção das escamas, as asas foram removidas do exemplar ainda seco e diafanizadas. Para os imaturos, exemplares foram trazidos de Rio Vermelho, São Bento do Sul, Santa Catarina, acondicionados em gaiolas de criação e casa de vegetação juntamente à planta hospedeira, foram realizados desenhos morfológicos com auxílio de câmara clara, fotografias dos diversos estágios e microscopia eletrônica de varredura de algumas estruturas. Dentre os resultados destacam-se: presença de banda transclipeal e de palpo maxilar rudimentar; união dos escleritos cervicais ventromedianamente e esclerito ventral situado anterior à união dos escleritos laterais; patágio membranoso; presença de almofada do basalar III, e sutura meral na coxa III. Seis ínstares larvais, e em todos, gregarismo durante o dia e dispersas para se alimentarem durante a noite. Duração total do ciclo: 73 dias. Palavras-chave: Bionomia; Ciclo de Vida; Citrus; Neotropical; Papilionoidea. 1
13 NATURAL HISTORY AND MORPHOLOGY OF Heraclides anchisiades capys (Hübner, [1809]) (LEPIDOPTERA : PAPILIONIDAE) ABSTRACT Papilionidae includes about 600 species, which are predominantly tropical butterflies. A subspecie which is very common in this family is Heraclides anchisiades capys, butterfly that has a dark coloring and feeds on Citrus spp. (Rutaceae). It has a relevant economical potential and it is easily raised in captivity, however, until now information about its biology is rare. The purpose of this study is to highlight the morphological structures of the adults and immatures of the specie, and the aspects related to the natural history, looking for a comparative focus with the other studied Papilionoidea and Hesperioidea. The structures of the adults were removed from the body, its appendages were isolated and boiled in KOH 10% to clarify and facilitate the removal of the scales, the wings were removed from the dry butterfly with and then clarified. For the immatures, samples were brought from Rio Vermelho, São Bento do Sul, Santa Catarina, kept in creation cages and greenhouse together with the host plant, morphological drawings with the help of light chamber, photos of several stages and scanning electronic microscopy of some structures were done. Among the results these are highlighted: presence of transclipeal band and rudimental maxillary palp; merging of the cervical sclerites ventromedially and ventral sclerite situated before the merging of the lateral sclerites; membranous patagia; presence of basalare III pad, and meral suture on the coxa III. Six larval stages were registered, and in all, gregarism during the day and spred to eat during the night. Total duration of the cycle: 73 days. Key words: Bionomy; Life cycle; Citrus; Neotropical; Papilionoidea. 2
14 CAPÍTULO I MORFOLOGIA DO ADULTO DE Heraclides anchisiades capys (Hübner, [1809]) (LEPIDOPTERA : PAPILIONIDAE) RESUMO A família Papilionidae inclui cerca de 600 espécies, as quais são predominantemente borboletas tropicais. Uma subespécie muito comum dentro dessa família é Heraclides anchisiades capys, borboleta de coloração escura que costuma se alimentar de Citrus spp. (Rutaceae). Apesar de comuns, nada se conhece em relação à morfologia desses insetos. O propósito deste estudo é evidenciar as estruturas morfológicas dos adultos da espécie, visando um enfoque comparativo com os demais Papilionoidea e Hesperioidea estudados. As estruturas foram removidas do corpo, entretanto, seus apêndices isolados e fervidos anteriormente em hidróxido de potássio (KOH) 10% para clarificar e facilitar a remoção das escamas, as asas foram removidas do exemplar ainda seco com o auxílio de pinça e tesoura e então diafanizadas. Posteriormente as estruturas foram desenhadas com auxílio de câmara clara e por fim interpretadas. Para maiores detalhamentos foram realizadas fotografias em microscópio eletrônico de varredura. Dentre os caracteres morfológicos encontrados destacam-se: Presença de banda transclipeal, sutura laterofacial em vista ventral próxima à metade da margem ocular de aspecto convexo, presença de palpo maxilar rudimentar. União dos escleritos cervicais ventromedianamente, esclerito ventral situado anterior à união dos escleritos laterais. Patágio membranoso, trocantin póstero-ventral ao episterno I unindo este à coxa protorácica. Anepisterno II fusionado ao processo pleural da asa II. Na asa anterior 3A presente partindo juntamente com a 2A e alcançando a margem interna da asa em seu terço proximal, 2A é a única veia anal presente na asa posterior. Presença de almofada do basalar III, e sutura meral na coxa III. Ausência de barra pós-espiracular no abdome e presença de superunco nos machos. Palavras - chave: Citrus; Neotropical; Papilioninae; Papilionoidea. 3
15 MORPHOLOGY OF THE ADULT OF Heraclides anchisiades capys (Hübner, [1809]) (LEPIDOPTERA : PAPILIONIDAE) ABSTRACT The Papilionidae includes about 600 species, which are predominantly tropical butterflies. A subspecie which is very common in this family is Heraclides anchisiades capys, butterfly that has a dark coloring and usually feeds on Citrus spp. (Rutaceae). Even though they are common, nothing is known about the morphology of these insects. The purpose of this study is to highlight the morphological structures of the adults of the specie, looking for a comparative focus with the other studied Papilionoidea and Hesperioidea. The structures were removed from the body, its appendages were isolated and boiled in KOH 10% to clarify and facilitate the removal of the scales, the wings were removed from the dry butterfly with the help of fine forceps and scissor and then clarified. Later the structures were drawn with the help of the light chamber and then interpreted. For further details, some pictures were taken on scanning electronic microscope. Among the morphological characters found, these are highlighted: Presence of transclipeal band, laterofacial suture in ventral view near the half of the ocular margin of convex aspect, presence of rudimental maxillary palp. Merging of the cervical sclerites ventromedially, ventral sclerite situated before the merging of the lateral sclerites. Membranous patagia, trochantin posteroventral to the episternum I gathering this to the prothoracic coxa. Anepisternum II fused to the pleural wing process II. On the forewing 3A present starting together with the 2A and reaching the inner margin of the wing in its proximal one-third, 2A is the only anal vein present on the hindwing. Presence of basalare III pad, and meral suture on the coxa III. Absence of post-espiracular bar on the abdome and presence of superuncus on males. Key words: Citrus; Neotropical; Papilioninae; Papilionoidea. 4
16 1. Introdução Lepidoptera é rica em espécies e ocorre em aproximadamente todas as regiões e com uma ampla variedade de habitats. A densa cobertura de escamas nas asas não apenas dá nome à ordem, mas forma a base para os padrões coloridos atrativos presentes em muitas espécies. A conspicuidade resultante da variação de Lepidoptera explica muito a atenção que estes insetos têm recebido (KRISTENSEN et al. 2007). Estimativas do número de espécies de lepidópteros variam consideravelmente. Segundo KRISTENSEN et al. (2007), existem cerca de espécies descritas. Os lepidópteros são praticamente iguais aos coleópteros em número de espécies fitófagas e pouco menos do que duas vezes o número de espécies fitófagas de hemípteros, a ordem com o número mais próximo das duas anteriores em relação às espécies que se alimentam de plantas (STRONG 1984, apud SCOBLE 1992). A importância ambiental e ecológica dos lepidópteros é grande pelo fato da maioria de suas larvas serem fitófagas e porque todos os estágios desses insetos são consumidos por predadores insetívoros ou parasitóides. A principal importância ecológica da alimentação no inseto adulto está relacionada com a polinização (SCOBLE 1992). Os lepidópteros se tornam de considerável importância para a humanidade quando as espécies desenvolvem o status de praga (SCOBLE 1992). A nocividade desses insetos resulta dos hábitos fitófagos das larvas, que muitas vezes causam prejuízos colossais, comportando-se como pragas das mais terríveis (COSTA LIMA 1945). Poucos insetos têm atraído mais atenção de biólogos do que borboletas da família Papilionidae. Sendo grandes e coloridas, são apreciadas por colecionadores amadores e têm sido objeto de pesquisas biológicas básicas (MILLER 1987). Segundo SCOBLE (1992), a família Papilionidae inclui cerca de 600 espécies, as quais são predominantemente borboletas tropicais. Geralmente de tamanho médio a grande, entre as mais espetaculares espécies de lepidópteros. A monofilia dessa família é sustentada por caracteres morfológicos como: presença de osmetério bifurcado no protórax das larvas; adulto com arólio e pulvilos dos pré- 5
17 tarsos reduzidos; asa anterior com veia 3A se estendendo livremente até a margem interna, ao passo que em outras espécies da mesma superfamília e nos demais lepidópteros, ela se funde com a veia 2A depois de uma curta distância ou falha ao alcançar a margem interna da asa. Uma subespécie comum dentro dessa família é Heraclides anchisiades capys (Hübner, [1809]), borboletas de coloração escura comumente vista por volta de bosques e campos de árvores cítricas, planta hospedeira das larvas (YOUNG et al. 1986) (Figs 1-4). Um resumo sobre a espécie é feito em COSTA LIMA (1968) como larva em folhas de Citrus spp. (bergamoteira e laranjeiras), coentrilho, guarantã e mamica de cadela. Parasitada por Pedinopelte gravenstrii (Guérin - Ménéville, 1826), Microcharops tibialis (Cresson, 1872) (Hymenoptera : Ichneumonidae); Apantela marquesi (Brêthes, 1924) (Hymenoptera : Braconidae); Spilochalcis sp. (Hymenoptera : Chalcididae). Predada por Apateticus mellipes (Bergroth, 1891), Alcaeorrhynchus grandis (Dallas, 1851), Oplomus catena (Drury, 1782) (Hemiptera : Pentatomidae), e distribuição registrada para: AM, PA, CE, RN, PE, PB, BA, MG, MT, ES, RJ, SP, SC, RS. Em LOPES et al. (1979), a larva dessa subespécie é descrita como sendo uma praga dos citros, principalmente na região nordeste. De acordo com BASTOS (1980), geralmente as larvas se encontram em grupos de 20 ou mais. Em árvores adultas não costumam causar grandes prejuízos, mas em árvores novas ou plantas que estejam sendo criadas em viveiros, podem causar sérios danos. Segundo JOHNSON & ROZYCKI (1986), Heraclides anchisiades capys (Hübner, [1809]) encontra-se exclusivamente na América do Sul, sem registros para as Américas Central e do Norte. Segundo GOMEZ (2002), a espécie possui grande potencial econômico e facilidade de criação em cativeiro, devido a seu número de posturas e taxa de sobrevivência. Contudo até os dias atuais são raras as informações sobre a biologia como já constatado por OLIVEIRA & URBAN (1977). De hábitos gregários tecem fios de seda permanecendo reunidas lado a lado, inclusive durante as ecdises, porém dispersam à noite para se alimentar. Quando bastante desenvolvidas, quase todas na quinta idade, passam a se reunir no tronco da planta hospedeira durante o dia (OLIVEIRA & URBAN 1977). 6
18 Uma vez fecundada, a fêmea procura a planta hospedeira Citrus spp. pousa na face ventral de uma folha, mantém as asas paralelas à mesma e as anteriores bastante inclinadas para trás, recurva o abdome para a face dorsal da folha, deposita um ovo e endireita o abdômen (OLIVEIRA & URBAN 1977). Na revisão bibliográfica, nenhuma informação foi encontrada sobre os caracteres morfológicos, estudos de comportamento nas diferentes fases do ciclo, mecanismos morfofuncionais de defesa e real importância econômica, visando dados quantitativos acerca dos prejuízos causados pela alimentação das larvas. Posição sistemática (LAMAS 2004): Heraclides anchisiades capys (Hübner, [1809]). PAPILIONOIDEA Papilionidae Papilioninae Papilionini Heraclides Hübner, [1809] anchisiades (Esper, 1788) (Papilio) a) anchisiades (Esper, 1788) (Papilio) b) capys (Hübner, [1809]) (Princeps) c) idaeus (Fabricius, 1793) (Papilio) d) lamasi K. S. Brown, 1994 e) philastrius (Fruhstorfer, 1915) (Papilio) As borboletas se encontram divididas em duas grandes superfamílias: Hesperioidea com cerca de espécies descritas e Papilionoidea com aproximadamente espécies, ambas as estimativas referem-se à região Neotropical (LAMAS 2004). Algumas das sinapomorfias que identificam os dois grupos podem ser encontradas na cabeça, como por exemplo, o espaço compreendido entre a base de inserção das antenas, enquanto naqueles este espaço é mensurável, nestes quase sempre a base dos alvéolos antenais estão muito próximas e em outras vezes chegam a se tocar medianamente. Também a forma da antena é fundamental já que em Hesperioidea a porção apical que é claviforme ou fusiforme curva-se em forma de gancho enquanto em Papilionoidea permanece reta. 7
19 O tórax em Lepidoptera é uma estrutura de grande importância na caracterização dos mesmos. ACKERY et al. (1999) relata a família Papilionidae como sendo provavelmente a mais bem documentada taxonomicamente entre todas as outras dentro de Lepidoptera. A monofilia da família é suportada por certo número de sinapomorfias únicas e universais, estando boa parte destas presentes no tórax (ACKERY 1984; SCOBLE 1992). Entretanto, a morfologia torácica de Lepidoptera tem sido estudada por poucos autores (SRIVASTAVA 1961), e de certa forma não utilizada em sistemas de classificação do grupo devido às estruturas deste tagma serem de difícil visualização, preparação e entendimento. Segundo SCOBLE (1992), o abdome constitui a parte mais obviamente segmentada dos insetos, o qual comporta a genitália, cujas estruturas são de grande interesse para a taxonomia do grupo. A estrutura básica das genitálias masculina e feminina é descrita, entretanto separadamente da sua função óbvia, os mecanismos precisos de suas partes particulares são relativamente pouco conhecidos. A morfologia da genitália feminina tem sido sistematicamente importante em categorias taxonômicas superiores, enquanto a genitália masculina é mais utilizada taxonomicamente em categorias inferiores como gênero e espécie. As estruturas pré-genitais também oferecem caracteres de importância, particularmente aqueles que formam a base do abdome e sua conexão com o tórax, esta região tem sido importante na reconstrução filogenética e muitas vezes negligenciada morfologicamente ao contrário dos caracteres genitais. Papilionidae é um grupo muito diverso e os caracteres morfológicos trabalhados são algumas vezes insatisfatórios. Alguns trabalhos focam as estruturas da genitália masculina e em particular a valva, justificando serem estas de importância morfológica ao se determinar as relações filogenéticas (HANCOCK 1983; MILLER 1987; TYLER et al. 1994). SRIVASTAVA (1965) trata a genitália masculina como uma estrutura que ainda leva a resultados controversos para a morfologia de Lepidoptera. Observa-se dessa forma que muitos trabalhos tratam a morfologia do abdome de Lepidoptera, entretanto o foco se dá apenas na genitália, enquanto que as estruturas pré-genitais não são abordadas (BANKS et al. 1976; CALLAGHAN 1997; KUZNETZOV & STEKOL NIKOV 2001; HALL & HARVEY 2002; HALL 2003; HALL & CALLAGHAN 2003). Para Papilionidae os trabalhos de MILLER (1987) e HANCOCK 8
20 (1983) também focam apenas as estruturas genitais. SRIVASTAVA (1965) abordou na maior parte de seu estudo o sistema muscular abdominal, justificando a negligência existente nos estudos de musculatura do abdome e genitália. 9
21 2. Objetivo Estudos com a morfologia comparada têm trazido para aprofundamentos, caracteres muitas vezes amplamente utilizados em determinadas famílias e completamente negligenciados em outras, supostamente porque exigem abundância de material para as dissecções e tempo para tal. Adultos de lepidópteros são o foco para estudos morfológicos, no entanto, com praticamente todos os caracteres levantados relacionados à genitália ou venação. Apesar de algumas espécies serem relativamente comuns ou até mesmo pragas, são escassas as bibliografias em relação à morfologia. Neste primeiro capítulo, este trabalho tem como objetivo evidenciar os componentes morfológicos do adulto de Heraclides anchisiades capys (Hübner, [1809]), visando um enfoque comparativo com os demais Papilionoidea e Hesperioidea já documentados. 10
22 3. Material e Métodos Exemplares utilizados neste estudo foram trazidos de Rio Vermelho, São Bento do Sul, Santa Catarina e outros coletados em jardins e parques de Curitiba e arredores, Paraná. Adultos são facilmente coletados quando estão visitando flores em busca de recursos alimentares. A cabeça e respectivos apêndices foram removidos do restante do corpo, entretanto fervidos anteriormente em hidróxido de potássio (KOH) 10% para clarificar e facilitar a remoção das escamas e posteriormente interpretados, desenhadas com auxílio de microscópio estereoscópico acoplado à câmara clara. As asas foram removidas do exemplar ainda seco com o auxílio de pinça e tesoura e então diafanizadas em placas de Petri, sendo primeiramente colocadas em álcool 70% e depois mergulhadas em Hipoclorito de Sódio (NaOCl) até a clarificação total, posteriormente submersas novamente em álcool 70% para a neutralização e então retiradas e secas em papel absorvente. Para o estudo da genitália, o abdome foi posicionado lateralmente e aberto longitudinalmente na região da pleura. Nos desenhos optou-se por pontuações nas regiões esclerotinizadas. A intensidade de pontos está diretamente relacionada à intensidade de esclerotinização das regiões. A realização de fotografias através de microscopia eletrônica de varredura seguiu procedimento padrão: primeiramente as peças passaram por desidratação através da imersão crescente em álcoois de diferentes concentrações 70%, 80%, 90% e 100%, 10 minutos em cada, sendo que no álcool absoluto as peças obtiveram um segundo banho de mais 10 minutos. Seguiu-se então o procedimento de ponto crítico, colagem do material em suportes específicos e metalização dos mesmos, para posterior realização das imagens. 3.1 Terminologia utilizada A falta de um trabalho universal de morfologia para Lepidoptera faz com que existam muitas divergências entre os termos empregados para as diversas estruturas do corpo. A terminologia que se emprega para as estruturas pertencentes à cabeça segue principalmente àquela de EHRLICH (1958a) no trabalho sobre Danaus plexippus Linnaeus, (Lepidoptera : Danaidae), com adaptações frente 11
23 à literatura clássica de morfologia (SNODGRASS 1935; MATSUDA 1965) e também outros trabalhos, alguns mais recentes (EHRLICH 1958b; CASAGRANDE 1979b; SORENSEN 1980; SCOBLE 1992; DUARTE et al. 2001; BIZARRO et al. 2003a; C. MIELKE et al. 2004a). Em relação às estruturas do tórax, a terminologia utilizada segue principalmente os trabalhos de SRIVASTAVA (1961, 1962) em seus estudos com Papilio demoleus Linnaeus, (Lepidoptera : Papilionidae), com modificações do autor frente aos trabalhos diversos de morfologia de Lepidoptera (SHEPARD 1930; MADDEN 1944; CASAGRANDE 1979c; SORENSEN 1980; BILOTTA 1994a; BIZARRO et al. 2003b; C. MIELKE et al. 2004b). Para as estruturas abdominais pré-genitais utiliza-se a terminologia usada por SRIVASTAVA (1965) em seu trabalho com Papilio demoleus Linnaeus, Em relação à genitália de ambos os sexos faz-se o uso principalmente da terminologia adotada por KLOTS (1956) e MILLER (1987) com adaptações que seguem (OITICICA FILHO 1946; EHRLICH 1958a, b, 1960; CASAGRANDE 1979d; SORENSEN 1980; BILOTTA 1994b; BIZARRO et al. 2003c, C. MIELKE et al. 2004c; DUARTE 2007), além de trabalhos clássicos de morfologia (SNODGRASS 1935; MATSUDA 1976). 3.2 Considerações gerais Segundo SNODGRASS (1935), o termo sutura refere-se a linhas de união entre áreas esclerotinizadas distintas. MATSUDA (1965) afirma ser difícil a distinção entre uma sutura verdadeira e um sulco externamente, concordando com o termo sutura antes definido. De acordo com SCOBLE (1992) e pelas mesmas justificativas de incertezas na identidade de sulco e sutura, o termo sulco é mais bem aplicado à cabeça, expondo então haver uma concordância geral quanto à homologia do termo dentro da ordem Lepidoptera e entre outras, confirmando SORENSEN (1980) que diz ser o termo sutura erroneamente aplicado na morfologia de Insecta, justificado pela falta de clareza na homologia do mesmo. Adota-se então para estes estudos morfológicos o termo sutura, seguindo essencialmente a terminologia adotada por EHRLICH (1958a). 12
24 3.3 Considerações sobre a cabeça MILLER (1971) chama de sutura laterofacial a linha que se estende da borda dos alvéolos antenais ventrolateralmente até o labro. De acordo com SORENSEN (1980) a homologia da sutura laterofacial ainda é incerta em Lepidoptera e requer estudos mais aprofundados, entretanto, propõe a existência de três tipos de sutura laterofacial. No tipo I esta surge junto à sutura do esclerito antenal seguindo paralelamente à margem anterior do olho até pouco acima da fóvea tentorial anterior onde toma um aspecto curvo ventromedianamente para alcançar esta. O tipo II assemelha-se ao tipo I, mas após a curva que alcança a fóvea tentorial anterior, a sutura laterofacial continua paralelamente à margem ocular, terminando ventralmente na cabeça. A sutura laterofacial do tipo III assemelha-se a do tipo II diferindo desta pelo fato de não alcançar a fóvea tentorial anterior. Para SNODGRASS (1935), esta mesma sutura denomina-se sutura frontal, o que para MATSUDA (1965) representa a sutura frontogenal. Segue-se então neste estudo a terminologia utilizada pela grande maioria dos autores (EHRLICH 1958a; MILLER 1971; SORENSEN 1980). De acordo com MILLER (1971), o rudimento mandibular é a estrutura articulada ou fundida com o frontoclípeo ou a área paraocular. SORENSEN (1980) por sua vez, diz que esta estrutura representa a mandíbula atrofiada nos Papilionoidea, a qual está fundida com a gena. Considera-se neste estudo a subgena como a região na qual o rudimento mandibular encontra-se fundido, concordando em posicionamento da estrutura com EHRLICH (1958a), que define o rudimento mandibular como uma pequena protuberância situada atrás e lateralmente ao pilífero. SNODGRASS (1935) denomina o vértice como sendo a região que ocupa o topo do crânio entre e após os olhos compostos. EHRLICH (1958a) diz que o mesmo se trata de uma região delimitada na porção anterior pela sutura transfrontal, lateralmente pelos alvéolos antenais e posteriormente pela sutura temporal. Apesar da inexistência de sutura transfrontal na espécie estudada, a descrição de EHRLICH (1958a) torna-se mais representativa para a determinação do vértice. Segundo EHRLICH (1958a) e SORENSEN (1980) a ponte tentorial é a estrutura que divide o forame magno. CASAGRANDE (1979b) e C. MIELKE et al. 13
25 (2004a) identificam esta como ponte pós-occipital. Segue-se o proposto por EHRLICH (1958a) considerando que esta estrutura faz parte do tentório. EHRLICH (1958a) define o occipício como a região lateral à sutura labial e sutura pós-occipital e que ocupa uma grande porção da superfície posterior da cápsula cefálica. Para SORENSEN (1980), a área entre os olhos, a sutura pósoccipital e a sutura hipostomal subgenal representam a pós-gena. A subgena separa-se póstero-ventralmente da pós-gena pela sutura hipostomal subgenal. A região posterior à subgena representa o occipício. Entretanto, nos macrolepidópteros a sutura occipital é questionável, a pós-gena e o occipício provavelmente se integraram formando na região posterior da cabeça a área pósgenal occipital. Pelo posicionamento adota-se a terminologia de SORENSEN (1980) para a área citada. 3.4 Considerações sobre a região cervical O cerviz é uma estreita região membranosa situada entre a cabeça e o tórax (SNODGRASS 1935). MATSUDA (1965) trata das divergências existentes entre diversos autores sobre a origem desta região, sem um consenso sobre a origem a partir da cabeça, tórax ou de ambos. BITSCH (1971) apud KRISTENSEN (2003) diz que morfologicamente o cerviz de Insecta possui fortes indícios de pertencer ao protórax, entretanto evidências embriológicas e de inervação sugerem a presença de um segmento labial compondo pelo menos parte da região dorsal deste esclerito. Devido à falta de maiores esclarecimentos sobre a origem desta estrutura, o trabalho trata a região cervical por questões práticas seqüencialmente à cabeça, independente de sua origem cefálica ou torácica. 3.5 Considerações sobre o tórax SRIVASTAVA (1961, 1962) trata o tórax como sendo dividido em duas partes distintas, o protórax como o primeiro segmento e o pterotórax como o segundo e que comporta os segmentos meso e metatorácicos. SNODGRASS (1935) cita que o tórax é uma região facilmente reconhecida, pois se trata da região que comporta as pernas e asas quando estas últimas se encontram presentes, o mesmo se divide em três partes distintas: protórax, mesotórax e metatórax, entretanto o autor realça que 14
26 os dois últimos segmentos também podem ser designados como pterotórax. O trabalho segue então a grande maioria dos autores que tratam o tórax em três partes distintas, sem menção ao termo pterotórax (EHRLICH 1958a, b; CASAGRANDE 1979c; SORENSEN 1980; SCOBLE 1992; C. MIELKE et al. 2004b). Os espiráculos pertencentes ao tórax causam divergências entres autores no que se refere à posição em relação aos segmentos. Alguns trabalhos (EHRLICH 1958a; CASAGRANDE 1979c) nomeiam os espiráculos como sendo primeiro e segundo pertencentes à membrana intersegmental do pro e mesotórax e da membrana intersegmental do meso e metatórax respectivamente. STEHR (1987) sugere a homologia entre as posições dos espiráculos no protórax e metatórax do adulto considerando a presença dos espiráculos nos mesmos segmentos torácicos dos imaturos. MADDEN (1944) trata os espiráculos como sendo meso e metatorácicos, entretanto sugere que o espiráculo do mesotórax teria migrado para a membrana pleural protorácica. Adota-se neste trabalho a denominação de primeiro e segundo espiráculos independente da posição que estes ocupam no tórax devido à falta de estudos mais aprofundados e que venham a esclarecer a origem destas estruturas. SRIVASTAVA (1962) descreve o acrotergito como sendo uma estreita placa marginal com um entalhe mediano no qual a região posterior da pós-zona protorácica se fixa ao mesotórax. MATSUDA (1970) trata a mesma região como uma estrutura marginal anterior a antecosta. De acordo com SORENSEN (1980), o acrotergito atravessa a margem anterior do mesonoto e ocorre como uma fina faixa marginal ao longo da sutura antecostal. Alguns autores não mencionam a presença deste esclerito (SHEPARD 1930; MADDEN 1944; EHRLICH 1958a; CASAGRANDE, 1979c; C. MIELKE et al. 2004b). Para o estudo em questão adota-se o termo acrotergito para a região supracitada, pela posição ocupada e concordância com o trabalho de SRIVASTAVA (1962). De acordo com MATSUDA (1970), o pronoto é composto tipicamente por uma placa dorsal mediana, a qual possui lateralmente áreas membranosas denominadas de patágio. Para SRIVASTAVA (1961) o pronoto é uma área membranosa exceto na região mediana que é esclerotinizada e dividida em duas partes distintas, a pró-zona anteriormente e a pós-zona posteriormente. Segundo o trabalho de EHRLICH (1958b) o patágio em Papilioninae tanto pode se apresentar esclerotinizado como membranoso. No presente estudo optou-se pela não indicação do patágio, devido a 15
27 não diferenciação deste em relação à área membranosa e para a terminologia das estruturas esclerotinizadas do protórax adota-se a proposta por SRIVASTAVA (1961). SNODGRASS (1935) chama de pré-episterno a subdivisão anterior do episterno, mas também faz uso do termo prepecto para a região, se a mesma estiver contínua com o pré-esterno. SRIVASTAVA (1962) considera o pré-episterno como sendo a região anterior ao episterno demarcada pela sutura pré-episternal. SHEPARD (1930) também utiliza o termo pré-episterno e o define como uma região anterior à sutura pré-episternal, sutura esta que se estende dorso-ventralmente desde a sutura anepisternal até o lobo esternal para se unir com a sutura pré-coxal. MATSUDA (1970) foca no fato da interpretação errônea do prepecto como sendo o pré-episterno por alguns autores. BROCK (1971) denomina de para-episterno a região de esclerotinização secundária próxima à margem anterior da placa infraepisternal basisternal, esta mesma região é denominada apenas como prepecto por SORENSEN (1980). Utiliza-se neste estudo devido à relação de posição que este esclerito ocupa e as demais estruturas próximas o termo préepisterno, seguindo assim a maioria dos autores que trabalharam estas estruturas (MADDEN 1944; EHRLICH 1958a; SRIVASTAVA 1962; CASAGRANDE 1979c; BILOTTA 1994a; CAMARGO et al. 2005). O basicoxito é uma estrutura geralmente em forma de estreita margem na base da coxa e próxima à sutura basicostal (SNODGRASS 1935). MATSUDA (1970) define o basicoxito como uma borda marginal demarcada externamente pela sutura basicostal na região terminal da coxa, e que forma uma área onde os côndilos da coxa se articulam com a pleura. A estrutura em questão observada não representa uma área esclerotinizada separada, e sim uma região de articulação entre os escleritos da eucoxa e mero ventralmente, e do catepisterno e epimero dorsalmente, sendo desta forma denominada neste estudo como articulação coxa-pleural, seguindo SRIVASTAVA (1962). 3.6 Considerações sobre o abdome e seus apêndices SRIVASTAVA (1965) denominou de látero-tergito o esclerito localizado ventralmente ao tergo I em ambos os lados do abdome, cuja superfície é fortemente sulcada. MADDEN (1944) denomina a mesma estrutura de esclerito pleural. Alguns 16
28 autores tratam este esclerito como barra tergo-pleural e sua projeção póstero-ventral de barra pós-espiracular (CASAGRANDE 1979d; BILOTTA 1994b; BIZARRO et al. 2003c; C. MIELKE et al. 2004c, DUARTE 2007). SORENSEN (1980) chama esta estrutura de esclerito secundário lateral. Devido à semelhança na conformação e ausência de demais projeções na estrutura, opta-se no presente estudo pelo uso do termo látero-tergito como citado por SRIVASTAVA (1965). Muitos autores tratam como vínculo o esclerito em forma de braço que se estende desde o saco localizado ventralmente até a projeção ântero-ventral do tegume (MADDEN 1944; KLOTS 1956; EHRLICH 1960; MUNROE 1961; MILLER 1987; HALL 1999; PENZ & DEVRIES 1999; KUZNETZOV & STEKOL NIKOV 2001). DUARTE (2007) denomina este mesmo esclerito como sendo o braço do tegume. O presente trabalho considera tal estrutura como sendo uma projeção ântero-dorsal do saco, sendo assim o termo adotado como mais adequado é o de braço do saco o que segue alguns autores que trataram desta mesma região (CASAGRANDE 1979d; BILOTTA 1994b; BIZARRO et al. 2003c; C. MIELKE et al. 2004c). O termo superunco utilizado neste estudo refere-se à extensão do tergo VIII em forma de garra que se curva em direção ventral, seguindo (KLOTS 1956; SRIVASTAVA 1965). Alguns autores denominam a mesma estrutura como pseudounco (HANCOCK 1983; MILLER 1987; ACKERY et al. 1999). MUNROE (1961) trata a estrutura como sendo um prolongamento posterior do oitavo tergo em formato de unco, sem nomear o mesmo. Fultura inferior é o termo adotado no estudo em questão para o esclerito em forma de U ventral ao edeago, pois se considera que este se trata de um sinônimo do termo juxta, antes tratado por diversos autores em diversos grupos de Lepidoptera (PIERCE 1909; MADDEN 1944; CASAGRANDE 1979d, PENZ & DEVRIES 2006; DUARTE 2007). PIERCE (1914) definiu a transtila como uma estrutura esclerotinizada na porção dorsal do diafragma, apresentando-se como uma barra transversal que se conecta ao ângulo dorso-proximal da valva, podendo apresentar sua região mediana algumas vezes ausente. A disposição encontrada neste estudo assemelha-se àquela citada por PIERCE (1914). SRIVASTAVA (1965) denominou o mesmo de esclerito sub-superuncal e MILLER (1987) o descreveu como esclerito dorso-lateral. 17
29 4. Resultados 4.1 Cabeça (Figs 5-30) Cabeça do tipo hipognata, com os olhos compostos ocupando a maior parte. Não apresenta ocelos e possui uma grande quantidade de escamas que recobrem toda a superfície da mesma. Na vista frontal os olhos compostos em destaque de forma globular e glabros, sendo sua largura superior à altura. Destes em direção ao frontoclípeo apresenta área paraocular entre a margem ocular e a sutura laterofacial, esta última que a separa da região frontoclipeal, esta sutura se apresenta mais próxima da margem ocular a partir da subgena. Frontoclípeo, uma grande área esclerotinizada em formato quadrangular entre os olhos compostos, pouco menor que a altura destes, ventralmente uma banda transclipeal marcada por estrias, está separada da subgena pela seção clipeo-genal da sutura laterofacial, na qual encontra-se contígua como uma invaginação do tegumento a fóvea tentorial anterior, dorsalmente o frontoclípeo não se encontra delimitado pela sutura transfrontal, não se tornando possível sua separação do vértice frontalmente. Labro juntamente com o pilífero, frontalmente se apresentam divididos em duas partes pela banda transclipeal, logo atrás deste em contato com a subgena tem-se o rudimento mandibular e ventralmente a gálea. Na região dorsal da cabeça a projeção pós-antenal com o quetosema destacado e posicionado posteriormente ao alvéolo antenal. Ventralmente a margem ocular forma um ângulo aberto e a sutura laterofacial, próxima à metade da margem ocular de aspecto convexo, forma uma curva que segue até as áreas do pilífero e do rudimento mandibular. A sutura hipostomal subgenal segue da região posterior da cabeça e encontra a sutura laterofacial na convexidade desta. Palpo maxilar rudimentar localizado na região distal do estipe com inúmeras cerdas. Ponte hipostomal separada do lábio pela sutura paralabial e da área pós-genal occipital pela sutura hipostomal subgenal. Em vista posterior, destacada após a margem ocular e em direção ao forame a área pós genal occipital ocupa a maior parte da área posterior da cabeça, tornando-se mais fracamente esclerotinizada em direção ao forame. A sutura hipostomal subgenal separa a área pós genal occipital da ponte hipostomal e termina junto à porção inferior do forame. Fóvea tentorial posterior, logo abaixo da 18
30 ponte tentorial junto à margem da porção inferior do forame. Forame magno superiormente delimitado pelo pós-occipício, medianamente dividido pela ponte tentorial, onde se encontra o côndilo occipital, lateralmente pela área pós-genal occipital e ventralmente pela ponte hipostomal. A projeção pós antenal e o quetosema são destacados e separados da área pós genal occipital através da sutura temporal que se estende ventralmente até pouco mais da metade da porção superior do forame, tomando a partir deste ponto um aspecto convexo até a sutura pós occipital. Em vista dorsal, a sutura laterofacial termina junto ao alvéolo antenal. Região frontoclipeal separada dorsalmente do vértice por diferenças de esclerotinização desde os alvéolos antenais e seguindo paralelamente à sutura laterofacial, uma vez que a sutura transfrontal é ausente. A projeção pós antenal e quetosema destacados e separados da área pós-genal occipital pela sutura temporal, que se estende até a região posterior da cabeça. Em vista lateral o olho composto se destaca ocupando a maior parte da cabeça. Frontoclípeo projetado. 4.2 Apêndices cefálicos Antena (Figs 11, 16-18) Clavada e sem carena, sem caracteres dimórficos e com pouca variação no número de artículos, ou seja, machos de 36 a 40, enquanto que as fêmeas de 38 a 41 flagelômeros. Em tamanho possui cerca de 8,2 vezes a largura do olho. Escapo, artículo proximal, é robusto e achatado dorso-ventralmente, com cerdas distalmente caracterizando região sensitiva. Pedicelo como um anel e metade do tamanho do escapo. Os demais formam o flagelo, sendo o primeiro alongado, do segundo ao sexto de tamanho reduzido e distalmente alongados como o primeiro, os sete últimos flagelômeros formam a clava, com o último de tamanho muito reduzido. 19
31 4.2.2 Aparelho bucal (Figs 6, 10, 12, 13, 19-27) Labio (Figs 6, 10, 12, 13, 19, 20) Representando uma estrutura pouco esclerotinizada, de forma triangular na região central da cavidade proboscidial, limitado lateralmente pelos estipes e ventralmente pela base dos palpos labiais, pela ponte hipostomal e separado desta pela sutura paralabial. Palpo labial quando visto lateralmente, apresenta-se voltado para cima e seu tamanho representa cerca de metade da largura do olho. Tri articulado, o artículo basal é o maior dos três, possui na face interna a mancha sensitiva de Reuter. O mediano é pouco maior que a metade do basal e o distal menor e com o órgão de Vom Rath na margem interna. Maxilas (Figs 6, 10, 19, 21-25) Na região anterior da fossa proboscidial formada pela gálea, estipe e cardo. Gálea não revestida por escamas, região apical com inúmeras papilas distribuídas de forma irregular, cerca de 6,9 vezes maior que a largura do olho. Estipe na base da gálea com formato de P inclinado e palpo maxilar rudimentar portador de inúmeras cerdas localizado na extremidade distal do estipe. Cardo reduzido de formato quadrangular. 4.3 Região cervical (Figs 14 e 15) Entre a cabeça e o tórax, reduzida quando comparada com as demais regiões do corpo. Lateralmente no centro da membrana o esclerito cervical em forma de T, articula anteriormente com a cabeça e posteriormente com a propleura. Se unem ventromedianamente, de forma mais evidente nos machos e fracamente esclerotinizada nas fêmeas. Ventralmente o órgão cervical como uma almofada revestida por inúmeras cerdas diferenciadas. Anterior à união dos braços dos escleritos está o esclerito ventral, retangular e ocupando o meio da região. 20
32 4.4 Tórax Protórax (Figs 31-34, 47, 48, 56) Menor entre os três segmentos torácicos e em sua maior parte formado por áreas membranosas. Pronoto constituído por duas regiões distintas, a pró-zona anteriormente e a pós-zona posteriormente, separadas pela sutura interzonal. A região pró-zonal possui formato de T, divida em braço transversal pró-zonal e braço longitudinal pró-zonal, separados pela sutura transversal pró-zonal. A sutura longitudinal pró-zonal atravessa o braço longitudinal pró-zonal desde a sutura transversal pró-zonal até a sutura interzonal. Pós-zona com formato de Y, possuindo lateralmente a sutura longitudinal pós-zonal em ambos os lados. Primeiro espiráculo póstero-ventralmente à pós-zona na pró-pleura e dorso-posterior ao episterno I. Em vista lateral, o episterno I anteriormente e ântero-ventralmente a este o basisterno I. Trocantin póstero-ventral ao episterno I unindo este à coxa protorácica, esta com cerdas diferenciadas na extremidade basal. Ventralmente o basisterno I com forma de T possuindo ânteromedianamente a ponte espinal-basisternal I. Lateralmente ao basisterno I, o episterno I, está separado deste pela sutura esterno-pleural. A furca, posterior ao basisterno I, articula na porção posterior com o espinasterno I que se apresenta dividido em braço transversal do espinasterno I e braço longitudinal do espinasterno I, este último de formato alargado e triangular posteriormente. O 1º espiráculo na margem da pró-pleura lateral ao episterno I Mesotórax (Figs 31, 33, 35, 36, 43, 44, 58, 59) Maior segmento torácico. Em vista dorsal o escuto II é mais estreito anteriormente, alargando-se em direção ao escutelo II. Acrotergito como uma fina banda anterior ao escuto II. Látero-anteriormente, a sutura escutal II separa o escuto II do suralar II e posteriormente o escutelo II separado do escuto II pela sutura escuto-escutelar II em formato de W, possuindo na extremidade posterior formato convexo em ambos os lados, unindo-se ântero-medianamente no escuto II. Posterior ao escutelo II, o pós-noto II triangular e o menor dos escleritos do mesotórax. 21
33 Lateralmente, o pré-escuto II anterior ao escuto II, e separado deste pela sutura préscuto-escutal, se projeta ventralmente formando o processo pré-alar II e posterior a este a subtégula alongada em forma de L invertido que alcança o processo pleural da asa II. Ventral à subtégula, o basalar II arredondado. Tégula dorso-lateralmente como um lóbulo móvel que articula na região posterior com a subtégula. Escuto II separado ventralmente do suralar II pela sutura escutal II e suralar II curvado em direção ao subalar II. Póstero-dorsal ao subalar II e como uma expansão póstero-ventral do escuto II, está o adnotal e, posterior a ele, o processo notal posterior da asa II como uma estreita estrutura em formato de C. Corda axilar II ventral à região posterior do escuto II e apoiada na extensão látero-ânterior do escutelo II. Ventral e posterior à corda axilar II, a ponte pós-alar se estende até o pós-noto II e, como uma estreita faixa, dorsal à corda axilar II surge o processo da ponte pós-alar. Região pleural dividida em duas regiões distintas: o episterno II proximal anterior e o epimero II distal posterior, separados pela sutura pleural II, sutura esta que se estende dorsal e longitudinalmente desde o processo pleural da asa II até a articulação coxa-pleural II ventralmente. Episterno II separado em anepisterno II e catepisterno II, situados dorsal e ventralmente respectivamente. O anepisterno II, uma pequena estrutura oval fusionado posteriormente ao processo pleural da asa II e separado ventralmente do catepisterno II pela sutura anepisternal II. Anterior ao catepisterno II e separado deste pela sutura pré-episternal II está o pré-episterno II e posterior ao mesmo o epimero II. Na porção ântero-dorsal do epimero II, uma dobra atinge o processo pleural da asa II e na região póstero-dorsal alcança a margem ventral da ponte pós-alar. Látero-posterior ao epimero II, o 2º espiráculo torácico na membrana intersegmental entre o meso e metatórax. Em vista ventral e anterior, o pré-episterno II localizado lateralmente ao braço longitudinal do espinasterno I. A sutura pré-episternal II separa posteriormente o préepisterno II do catepisterno II e corre transversalmente em ambos os lados, unindose ântero-medianamente para formar o discrime II, que se estende longitudinalmente até a fóvea apofisial II, formando uma invaginação do esterno II. Ainda anterior à união da sutura pré-episternal II, uma porção não invaginada do esterno II em formato triangular. 22
34 4.4.3 Metatórax (Figs 31, 33, 35, 36, 49, 57) Segundo maior esclerito torácico. Dorsalmente dividido anteriormente em escuto III e posteriormente em escutelo III, separados pela sutura escuto-escutelar III. Anterior ao escuto III, uma área membranosa que o separa do pós-noto II. Escuto III mais estreito medianamente, alargando-se gradativamente em direção às margens laterais, formando duas áreas de aspecto subtriangulares. Enquanto o escutelo III, de formato triangular e menor, situa-se posteriormente. Lateralmente o escuto III sub-quadrangular, separa-se ventralmente do suralar III pela sutura escutal III, posteriormente pela sutura escuto-escutelar III do escutelo III sub-retangular, este desce dorso-ventralmente até a região posterior do processo notal posterior da asa III. Ventral ao escutelo III, apoiada a corda axilar III e póstero-ventralmente a esta última, o pós-noto III não visível dorsalmente. Subalar III, uma estreita estrutura situado aproximadamente no centro da membrana pleural. Região pleural III com conformação semelhante à região pleural II, sendo que ântero-ventralmente ao catepisterno III situa-se o basisterno III, não observado no mesotórax. Ântero-dorsalmente ao catepisterno III, o basalar III arredondado, possui uma estrutura sobreposta na margem ventral denominada almofada do basalar III, membranosa e com inúmeras cerdas dorsalmente. O epimero III se estende dorsoposteriormente até a porção anterior do pós-noto III. Em vista ventral, o discrime III se estende médio-longitudinalmente desde a margem anterior do metatórax até a fóvea apofisial III posteriormente. O basisterno III, lateral ao discrime III, como uma placa sub-retangular. 4.5 Apêndices torácicos Pernas (Figs 37-42) Apresentando um par de pernas por segmento torácico, cada uma composta de coxa, trocânter, fêmur, tíbia e tarso. Perna protorácica a menor. Coxa I alongada, mais larga na extremidade basal e se estreitando gradativamente em direção distal, articula com a pleura através de um pequeno esclerito em forma de taça, o trocantin. Sutura coxal I se estende desde a base da coxa I até aproximadamente um quinto do comprimento da mesma. Trocânter sub-retangular, o menor dentre os 23
35 escleritos da perna. Tíbia I apresenta medianamente na superfície posterior uma estrutura foliar, a epífise. Tarso I composto de cinco tarsômeros, sendo o proximal o maior deles, possuindo aproximadamente quatro vezes o comprimento dos demais, que possuem comprimentos semelhantes. Espinhos são visualizados em todo comprimento da tíbia e tarso, distribuídos de forma irregular na superfície ventral da tíbia e de forma regular formando duas fileiras de espinhos na superfície ventral dos tarsômeros. No distitarso uma formação distal de cerdas diferenciadas situadas ventralmente entre os espinhos. Garras tarsais alongadas, curvas e intermediadas basal-ventralmente na membrana pela placa ungüitratora, de formato quadrangular. Pernas meso e metatorácica apresentam conformação semelhante. Coxa II dividida longitudinalmente em duas partes através da sutura coxal II, a eucoxa II anterior e o mero II posterior. Coxa III semelhante à coxa II, entretanto o mero III comporta uma sutura transversal que o divide em duas regiões, a sutura meral. Um par de esporões se encontra articulado à extremidade distal interna das tíbias meso e metatorácica Asas e escleritos axilares (Figs 31-34) Asas anteriores triangulares em ambos os sexos. Margem externa com os terços superior e inferior de aspecto convexo, enquanto o terço médio é côncavo na altura de M 2 e M 3, característica esta mais pronunciada nas fêmeas. Subcostal (Sc) parte da região axilar mais espessa na base, terminando junto à margem costal em seu terço distal. Radial (R) surge junta e paralela à Sc e ao atingir o terço distal da célula discal bifurca-se em R 1 e Rs e posteriormente, esta se divide em R 2, R 3, R 4 e R 5, R 3 terminando no ápice e R 4 e R 5 na margem externa. Margens costal e externa se encontram formando o ápice da asa, pontiagudo nos machos e mais arredondado nas fêmeas. Nos machos, M 1, M 2 e M 3 partem separadas do ápice da célula discal, M 2 mais próxima de M 1 e M 3 mais afastada e curvada para baixo em direção à margem externa. Ainda nos machos dci é ligeiramente maior que dcm. Nas fêmeas as veias dcm e dci de igual tamanho e M 1, M 2 e M 3 eqüidistantes desde a base até a margem externa. CuA 1 e CuA 2 partem separadas da célula discal por uma distância equivalente a extensão de m-cu, sendo que nas fêmeas CuA 2 apresenta-se curvada para cima, o que não ocorre nos machos. CuP curta partindo do terço anterior da Cubital e alcançando o terço anterior de 2A. Veias 2A e 3A partem da região axilar 24
36 separadas da célula discal, 2A atinge o ângulo anal da asa, mais próxima da margem interna nos machos e mais distante nas fêmeas. Veia 3A curva em direção à margem interna e alcança esta no terço proximal. Asas posteriores convexas nas margens costal e externa e côncavas na margem interna. Veia Sc + R 1 desde a região axilar até o ápice da asa. Veia humeral (h) curva em direção ao ápice da asa e surge junto a Sc + R 1 na base destas, onde ambas se separam ligeiramente para formar a célula basal. Veia dcs maior que dcm e esta maior que dci. CuA 1 e CuA 2 partem separadamente da célula discal por uma distância ligeiramente maior que o comprimento de m-cu. Veia 2A, a única anal, termina no ângulo anal da asa. Escleritos axilares representam a região que conecta a asa ao tórax, constituída por vários escleritos de formatos irregulares. Na asa anterior, láteroposteriormente ao escuto II, a placa pós-alar possui anteriormente o processo notal mediano da asa II e ântero-lateralmente o processo notal posterior da asa II, o primeiro deles se articula ao primeiro axilar e o segundo ao terceiro axilar, neste último a articulação se dá através de uma dobra da membrana. Terceiro axilar articula látero-distalmente às regiões basais da Cubital e da Anal. Segundo esclerito axilar distal ao primeiro e terceiro, possui formato trilobado, com o lobo anterior articulado ao 1º axilar, o mediano ao 3º axilar e o posterior aos escleritos basais da Sc e R. Distalmente ao 2º axilar articula-se o processo basal subcostal II. Nas asas posteriores o processo notal posterior se encontra láteroposteriormente ao escutelo III e articula distalmente com o 3º axilar, este último látero-posteriormente articula com os escleritos basais da veia 2A e distalmente com o 2º axilar. O 1º axilar articula com o 2º axilar lateralmente e este último distalmente com o processo basal subcostal III. 4.6 Abdome Estruturas pré-genitais (Figs 60-63) Abdome composto por 10 segmentos, sendo que os dois últimos são modificados nos machos dando origem as estruturas da genitália, enquanto que nas fêmeas os três últimos. Cada segmento pré-genital é composto por um tergo dorsalmente e um esterno ventralmente, ambos esclerotinizados e conectados na 25
37 região da pleura, sendo esta uma área membranosa que apresenta em cada segmento o respectivo espiráculo de forma elíptica. As fêmeas apresentam o abdome mais alargado dorso-ventralmente em relação aos machos. Em ambos os sexos, o primeiro segmento apresenta modificações em seus escleritos, relativas à junção com a região posterior do metatórax. Tergo I com terço anterior membranoso, sendo o restante esclerotinizado e retangular. Ventralmente a este último, o látero-tergito, como um esclerito estreito e projetado anteriormente, nos machos de forma subtriangular e nas fêmeas subretangular com pequena projeção póstero-ventral. Esterno I anterior ao espiráculo I, delgado e projetado anteriormente em direção à região ântero-mediana do abdome e posteriormente articulado a extremidade ântero-dorsal do esterno II. Tergo VII nos machos mais estreito em relação aos demais e tergo VIII com projeção delgada posterior formando o superunco. Nas fêmeas, tergo VIII mais estreito em relação aos demais. Esterno VIII retangular e o último observado nos machos antes dos elementos genitais, enquanto nas fêmeas o esterno VII é o último e ovalado Genitália masculina (Figs 64-68) Dorsalmente limitada pelo oitavo tergo, projetado mediano-posteriormente em uma estrutura em gancho curvada dorso-ventralmente, denominada de superunco. Tegume representando o tergo IX, médio-ventral ao tergo VIII, fundido anteriormente ao unco, este último uma modificação do tergo X, triangular com projeção distal curvada ventralmente. Gnato articulado à margem posterior do tegume, modificação do décimo esterno, como um esclerito ventral, projetado em direção caudal e com cerdas dorsais. Transtila reduzida a um par de estruturas delgadas, com região mediana ausente, e articulada no ângulo dorso-proximal da valva. Fultura inferior, estreita estrutura esclerotinizada em forma de U, situada ventralmente dando sustentação ao edeago e visível apenas em vista posterior. A genitália é limitada em sua porção ventral pelo saco, modificação do nono esterno com o braço do saco projetado ântero-dorsalmente como uma fina banda esclerotinizada que atinge o braço ventral do tegume, formando um arco. Valva achatada médio-lateralmente, formando um par de placas articuladas à transtila na região da costa e aos braços ventrais do tegume e dorsais do saco na 26
38 região do sáculo. A valva apresenta em sua porção central a harpe, com um grupo de cerdas diferenciadas em posição ântero-dorsal, borda posterior denteada e fortemente esclerotinizada em relação às demais regiões. Edeago cilíndrico, com terço anterior fortemente esclerotinizado e terço posterior com fenda distal lateralmente. Bulbo ejaculatório terminal, sendo o local de passagem do duto ejaculatório. Distalmente observa-se a vesica sem cornuto que permanece retraída em situação de repouso Genitália feminina (Figs 69 e 70) Constituída pelos oitavo, nono e décimo segmentos do abdome modificados. Tergo VIII com projeção ântero-ventral em ambos os lados, formando as apófises anteriores, e tergos IX + X formando as papilas anais, sendo estas duas, placas verticais póstero-medianamente localizadas no abdome, com esclerotinização parcial e inúmeras cerdas distribuídas na superfície. Apófises posteriores como um par de projeções ântero-laterais das papilas anais. Esterno VIII representando por duas placas genitais, anteriormente a lamela ante-vaginal e posteriormente a lamela pós-vaginal, entre estas se abre o óstio da bolsa. Bolsa copuladora formada pelo óstio, duto e corpo da bolsa. O duto da bolsa se inicia no óstio, e se insere na bolsa, esta membranosa e de forma saculiforme, apresentando apenas um signo ventro-lateralmente esquerdo que ocupa todo seu comprimento, a bolsa apresenta aproximadamente 2,3 vezes o comprimento do duto. 27
39 5. Discussão 5.1 Cabeça Entre machos e fêmeas da espécie estudada não houve diferenças significativas em relação às estruturas da cabeça. Frontalmente a sutura clípeo-labral presente nos demais Papilionoidea (EHRLICH 1958a; CASAGRANDE 1979b; SORENSEN 1980; BIZARRO et al. 2003a; C. MIELKE et al. 2004a) e em Hesperioidea (MILLER 1971), é em Heraclides anchisiades capys ausente como também para Papilio demoleus Linnaeus, 1758 conforme o observado por (SRIVASTAVA 1957, apud SORENSEN 1980). A banda transclipeal presente, assim como em Hesperioidea MILLER (1971) e poucos Papilionoidea (EHRLICH 1958a; BIZARRO et al. 2003a). EHRLICH (1958a) define esta banda como sendo uma região de esclerotinização mais intensa ocupando o quinto inferior do esclerito frontoclipeal. Na maioria dos outros Papilionoidea esta estrutura parece estar ausente (CASAGRANDE 1979b; SORENSEN 1980; DUARTE et al. 2001; C. MIELKE et al. 2004a). Os olhos compostos se destacam frontalmente com largura superior à altura, diferentemente do que ocorre em Hesperioidea MILLER (1971), também não se apresentando dividido em duas áreas distintas, característica esta particular dos Hesperioidea. Região frontoclipeal não separada do vértice pela sutura transfrontal, sutura esta presente em outros Papilionoidea como Nymphalidae (EHRLICH 1958a; BIZARRO et al. 2003a; C. MIELKE et al. 2004a) e Lycaenidae (SORENSEN 1980; DUARTE et al. 2001). A sutura laterofacial em vista ventral próxima à metade da margem ocular de aspecto convexo, formando uma curva que atinge as áreas do pilífero e rudimento mandibular, esta característica não observada em outros Papilionoidea ou em Hesperioidea. Entre os apêndices, as antenas tradicionalmente usadas para diferenciar os Papilionidae de Nymphalidae principalmente pelas carenas presentes nestas, não apresentam caracteres dimórficos apenas com pequena variação no número de flagelômeros. Palpo maxilar rudimentar presente, esta característica não observada até a presente data nos demais Papilionoidea, entretanto em Hesperioidea tal estrutura é 28
40 evidenciada por MILLER (1971). Diferentemente do que ocorre com muitos outros Papilionoidea (EHRLICH 1958a; CASAGRANDE 1979b; SORENSEN 1980; BIZARRO et al. 2003a) e em Hesperioidea MILLER (1971), o palpo labial apresenta o artículo basal como sendo o de maior comprimento. Região cervical formada por duas áreas distintas, os escleritos cervicais laterais que se unem ventromedianamente e um esclerito ventral situado anteriormente à união dos escleritos cervicais laterais quando observado ventralmente. SRIVASTAVA (1961) observou esta mesma disposição dos escleritos cervicais trabalhando com Papilio demoleus L. (Lepidoptera : Papilionidae). Esta característica não foi identificada até o momento em outros Papilionoidea ou em Hesperioidea, onde os braços ventrais não se fundem. MADDEN (1944), no entanto, em seu trabalho sobre Manduca sexta Linnaeus, 1763 (Lepidoptera : Sphingidae), refere-se à união dos braços ventrais dos escleritos cervicais laterais em posição medio-ventral, como o observado no presente estudo, mas faltam estudos morfológicos em outros grupos de noturnas para que possamos ampliar as comparações. 5.2 Tórax Machos e fêmeas não apresentam diferenças significativas em relação às estruturas torácicas, com exceção do formato das asas. O pronoto apresenta patágio membranoso como observado por EHRLICH (1958b) não se tornando possível sua delimitação na área pronotal, assim como em Papilio demoleus L., (SRIVASTAVA 1961). Em outros Papilionoidea, o patágio é evidente e se apresenta esclerotinizado ântero-dorsalmente no pronoto (EHRLICH 1958a; CASAGRANDE 1979c; SORENSEN 1980; BILOTTA 1994a; C. MIELKE et al. 2004b). O trocantin, como um pequeno esclerito distal à coxa I, esta presente em alguns grupos de Papilionoidea (SRIVASTAVA 1961; C. MIELKE et al. 2004b), e em outros (EHRLICH 1958a; CASAGRANDE 1979c; SORENSEN 1980; BIZARRO et al. 2003b) ausente, ou não citado. Na asa anterior, 3A presente de origem comum com 2A e alcançando a margem interna da asa em seu terço proximal. Em alguns grupos de Papilionoidea, 29
41 3A é ausente (EHRLICH 1958a; CASAGRANDE 1979c; BILOTTA 1994a; C. MIELKE et al. 2004b). Em outros representantes desta mesma superfamília a mesma aparece na asa anterior, entretanto curvada para cima e termina ao alcançar o terço proximal de 2A (SORENSEN 1980; BIZARRO et al. 2003b; DUARTE 2007). Em Hesperioidea EHRLICH (1960) estudou Epargyreus clarus (Cramer, 1775) (Lepidoptera - Hesperiidae) e identificou a presença de 3A, curvada para baixo e falha ao alcançar a margem interna da asa. SRIVASTAVA (1962) observou uma disposição dos escleritos axilares da asa anterior diferente da verificada no presente estudo. Em Papilio demoleus L., o 1º axilar articula anteriormente ao suralar II e o 2º axilar anteriormente ao processo notal mediano da asa II. Em Heraclides anchisiades capys, o 1º axilar articula com o processo notal mediano da asa II e o 2º axilar, distalmente ao primeiro não faz qualquer ligação com o suralar II. Uma sutura marginal observada por SRIVASTAVA (1962), paralela à margem anterior da suralar II em Papilio demoleus não é visualizada na espécie estudada. Veia 2A é a única anal presente na asa posterior assim como em Papilio demoleus SRIVASTAVA (1962). Em outros Papilionoidea e Hesperioidea já documentados até a presente data, a veia 3A também é encontrada na asa posterior. Na coxa metatorácica, o mero é dividido transversalmente em duas regiões pela sutura meral, sutura esta já descrita para Papilionidae EHRLICH (1958b) e como observada em Papilio demoleus por SRIVASTAVA (1962). Não há registros desta estrutura para outros Papilionoidea e Hesperioidea. Tíbia metatorácica com apenas um par de esporões tibiais assim como os demais Papilionoidea e contrário ao verificado em muitos Hesperioidea, com dois pares de esporões como citado por (EHRLICH 1960). Ântero-dorsalmente ao catepisterno III, ventralmente ao basalar III e apoiando este último, a almofada do basalar III, como uma estrutura membranosa com cerdas distribuídas na superfície dorsal. Em Lycaenidae (SORENSEN 1980; DUARTE 2007) a almofada do basalar III também é citada, como em Papilio demoleus por SRIVASTAVA (1962) e por MADDEN (1944) para Sphingidae. Esta estrutura é completamente ausente nos demais Papilionoidea e Hesperioidea até então estudados. 30
42 5.3 Abdome EHRLICH (1958b) relata a ausência da barra pós-espiracular em Papilionidae, o que se corrobora no presente estudo, entretanto, em relação à barra préespiracular, o mesmo autor diz ser ausente apenas em Pieridae e em nenhum momento comenta sobre a condição desta mesma barra nos distintos grupos. No presente estudo, a barra pré-espiracular encontra-se indistinta, sendo que na região somente o primeiro esterno é visualizado, assim como encontrado por SRIVASTAVA (1965). BILOTTA (1994b) não menciona a barra pré-espiracular em Morphinae (Nymphalidae), entretanto o esclerito se encontra em outros ninfalídeos (EHRLICH 1958a; CASAGRANDE 1979d; BIZARRO et al. 2003c; C. MIELKE et al. 2004c). Nos demais Papilionoidea e Hesperioidea estudados morfologicamente até o momento a barra pré-espiracular está presente. O superunco é até o presente estudo, uma sinapomorfia apenas para Papilioninae, entretanto MILLER (1987) observou estrutura similar em Colias philodice Godart, 1819 (Lepidoptera Pieridae). Este esclerito aparece nos diversos trabalhos de sistemática dentro da família, sendo apresentado sob formas singulares, inclusive na espécie em estudo, em geral de formas totalmente distintas para cada espécie dentro de Papilionidae (SRIVASTAVA 1965; HANCOCK 1983; MILLER 1987; TYLER et al. 1994), o mesmo acontece com outras estruturas da genitália masculina de grande importância taxonômica, pelas particularidades apresentadas nestes trabalhos para cada espécie, como são os casos da transtila, gnato, fultura inferior e valva. A projeção do saco na espécie estudada é posterior, desta forma contrária aos demais Papilionoidea já estudados onde a mesma é anterior (CASAGRANDE 1979d; BILOTTA 1994b; BIZARRO et al. 2003c; C. MIELKE et al. 2004c). Nas fêmeas, a bolsa copuladora é estruturalmente bastante distinta entre as espécies de Papilionidae e os demais Papilionoidea e Hesperioidea. Forma, número, posição e tamanho dos signos da bolsa nas diferentes espécies observadas nos estudos desta estrutura dentro da família (MUNROE 1961; HANCOCK 1983; MILLER 1987) e em outros grupos (EHRLICH 1960; CASAGRANDE 1979d; HALL 2001; PENZ & DEVRIES 2006; DUARTE 2007), mostram que as mesmas são de significativa importância taxonômica. 31

43 Figs 1-4. Heraclides anchisiades capys. Figs Macho: 1 - Vista dorsal. 2 - Vista ventral. Figs Fêmea: 3 - Vista dorsal. 4 - Vista ventral. 32
44 5 6 7 Figs 5-7. Heraclides anchisiades capys - Cabeça: 5 - Vista frontal. 6 - Vista ventral. 7 - Detalhe da vista ventral da cabeça. 33
45 8 9 Figs 8 e 9. Heraclides anchisiades capys - Cabeça: 8 - Vista posterior. 9 - Vista dorsal. 34
46 10 Fig.10. Heraclides anchisiades capys - Cabeça: Vista lateral. 35
47 Figs Heraclides anchisiades capys Antena Palpo labial: 12 - Vista externa Vista interna. 36
48 14 15 Figs 14 e 15. Heraclides anchisiades capys - Região cervical: 14 - Vista lateral Vista ventral. 37



49 Figs Heraclides anchisiades capys - Antena: 16 - Escapo Detalhe da área sensitiva Artículo distal da antena. 38







50 Figs Heraclides anchisiades capys - Cabeça: 19 - Vista ventral da cabeça Artículo distal do palpo Cardo Palpo maxilar rudimentar com cerdas diferenciadas Ápice da gálea. 39





51 Figs Heraclides anchisiades capys - Cabeça: Região do pilífero e rudimento mandibular Fóvea tentorial anterior Fóvea tentorial posterior Quetosema. 40
52 31 32 Figs 31 e 32. Heraclides anchisiades capys - Tórax: 31 - Vista dorsal Protórax em vista dorsal. 41
53 33 Fig. 33. Heraclides anchisiades capys - Tórax em vista lateral. 42
54 34 35 Figs 34 e 35. Heraclides anchisiades capys - Tórax: 34 - Protórax vista ventral Mesotórax e Metatórax vista ventral. 43
55 36 Fig. 36. Heraclides anchisiades capys - Escleritos axilares vista dorsal. 44
56 Figs Heraclides anchisiades capys - Perna protorácica: 38 - Detalhe da epífise Detalhe do último tarsômero. 45
57 40 41 Figs 40 e 41. Heraclides anchisiades capys - Perna mesotorácica: 41 - Detalhe da região do esporão tibial. 46
58 42 Fig. 42. Heraclides anchisiades capys - Perna metatorácica. 47
59 43 44 Figs 43 e 44. Heraclides anchisiades capys - Tégula: 43 - Vista lateral anterior Vista lateral posterior. 48
60 45 Fig. 45. Heraclides anchisiades capys - Asas anterior e posterior - macho. 49
61 46 Fig. 46. Heraclides anchisiades capys - Asas anterior e posterior - fêmea. 50



62 Figs Heraclides anchisiades capys - Tórax: Trocantin e extremidade basal da coxa I com cerdas diferenciadas Articulação coxa-pleural. Cx - coxa. Tr - trocantin. 51






63 Figs Heraclides anchisiades capys - Perna protorácica: Epífise tibial Distitarso vista lateral Distitarso vista ventral. 52




64 Figs Heraclides anchisiades capys - Tórax: 56 - Basisterno I Almofada do basalar III Tégula vista geral Tégula vista da região de cerdas. 53
65 60 Fig. 60. Heraclides anchisiades capys - Abdome do macho em vista lateral. 54
66 61 Fig. 61. Heraclides anchisiades capys - Abdome do macho em vista ventral. 55
67 62 Fig. 62. Heraclides anchisiades capys - Abdome da fêmea em vista lateral. 56
68 63 Fig. 63. Heraclides anchisiades capys - Abdome da fêmea em vista ventral. 57
69 64 65 Figs 64 e 65. Heraclides anchisiades capys - Genitália masculina: 64 - Vista lateral Vista posterior. 58
70 Figs Heraclides anchisiades capys - Genitália masculina: 66 - Valva Edeago em vista lateral anterior Edeago em vista lateral posterior. 59
71 69 Fig. 69. Heraclides anchisiades capys - Genitália feminina vista ventral. 60
72 70 Fig. 70. Heraclides anchisiades capys - Genitália feminina em perfil. 61
73 6. Referências Bibliográficas ACKERY, P, R; R. DE JONG & R. J. VANE-WRIGHT The Butterflies Hydeloidea, Hesperioidea and Papilionoidea, p In: P. N. KRISTENSEN (Ed.). Band/Volume IV Arthropoda: Insecta. Teilband / Part 35.Lepidoptera moths and butterflies: evolution, systematics and biogeography. Vol. 1, X+491p. In: M. FISCHER (Ed.). Handbook of Zoology. Berlin, New York, Walter de Gruyter. ACKERY, P. R Systematic and faunistic studies on butterflies, pp.9 21, 1 fig. In: VANE-WRIGHT, R. I. & P. R. ACKERY (Eds.). The Biology of Butterflies, Academic Press, 429 pp. BANKS, H. J; J. D. HOLLOWAY & H. S. BARLOW A revision of the genus Ptychandra (Lepidoptera Nymphalidae). Bulletin of the British Museum (Natural History) Entomology, 32: BASTOS, J. A. M Ensaio preliminar de controle da lagarta dos citros, Papilio anchisiades capys (Hübner, 1809) com inseticidas orgânicos sintéticos em laboratório. Fitossanidade, 1980, publ. 1981, 4 (1): 6-7. BILOTTA, I Morfologia comparada do tórax das espécies sulbrasileiras de Morphinae (Lepidoptera Nymphalidae). Revista Brasileira de Zoologia, 11 (4): BILOTTA, I Morfologia comparada do abdome das espécies sulbrasileiras de Morphinae (Lepidoptera Nymphalidae). Revista Brasileira de Zoologia, 11 (4): BIZARRO, J. M. S; M. M. CASAGRANDE & O. H. H. MIELKE Morfologia externa de Thyridia psidii cetoides (Rosemberg & Talbot). I. Cabeça e apêndices (Lepidoptera, Nymphalidae, Ithomiinae). Revista Brasileira de Zoologia, 20 (2):
74 BIZARRO, J. M. S; M. M. CASAGRANDE & O. H. H. MIELKE Morfologia externa de Thyridia psidii cetoides (Rosemberg & Talbot) II. Tórax e appendices (Lepidoptera, Nymphalidae, Ithomiinae). Revista Brasileira de Zoologia, 20 (3) BIZARRO, J. M. S; M. M. CASAGRANDE & O. H. H. MIELKE Morfologia externa de Thyridia psidii cetoides (Rosenberg & Talbot) (Lepidoptera, Nymphalidae, Ithomiinae). III. Abdome e apêndices. Revista Brasileira de Zoologia, 20 (4): BROCK, J. P A contribution towards an understanding of the morphology and phylogeny of the Ditrisian Lepidoptera. Journal of Natural History, 5: CALLAGHAN, C. J A review of the genus Panara Doubleday, 1847 (Riodinidae) in southeast Brazil, with a description of two new subspecies. Journal of Research on the Lepidoptera, 34: CAMARGO, A. J. A; M. M. CASAGRANDE & O. H. H. MIELKE Morfologia externa do adulto de Almeidaia aidae Mielke & Casagrande (Lepidoptera, Saturniidae, Arsnurinae, Almeidaiinae). II. Tórax e apêndices. Revista Brasileira de Zoologia, 22 (4): CASAGRANDE, M. M Sobre Caligo beltrao (Illiger). II: Morfologia externa da cabeça do adulto (Lepidóptera, Satyridae, Brassolinae). Revista Brasileira de Biologia, 39 (1): CASAGRANDE, M. M Sobre Caligo beltrao (Illiger). III: Morfologia externa do adulto tórax. (Lepidoptera, Satyridae, Brassolinae). Revista Brasileira de Biologia, 39 (2): CASAGRANDE, M. M Sobre Caligo beltrao (Illiger). IV: Morfologia externa do adulto - abdome. (Lepidoptera, Satyridae, Brassolinae). Revista Brasileira de Biologia, 39 (3):
75 COSTA LIMA, A. M Quarto catálogo dos insetos que vivem nas plantas do Brasil seus parasitos e predadores. Parte II 1º tomo. Min. Agric., Rio de Janeiro. COSTA LIMA, A. M Insetos do Brasil. Lepidópteros 1ª parte, 5º tomo, cap. XXVIII., Escola Nacional de Agronomia, série didática nº 7, 379p. DUARTE, M; M. M. CASAGRANDE & O. H. H. MIELKE Morfologia externa do adulto de Hemiargus hanno (Stoll) (Lepidoptera, Lycaenidae, Polyommatinae, Polyommatini). I. Cabeça. Revista Brasileira de Zoologia, 18 (1): DUARTE, M Morfologia externa do adulto de Hemiargus hanno (Stoll) (Lepidoptera, Lycaenidae, Polyommatinae, Polymmatini) II. Região cervical, tórax e abdome. Iheringia, Série Zoologia, 97(2): EHRLICH, P. R The integumental anatomy of the monarch butterfly Danaus plexippus L., (Lepidoptera: Danaidae). The University of Kansas Science Bulletin, 38: EHRLICH, P. R The comparative morphology, phylogeny, and the higher classification of the butterflies (Lepidoptera: Papilionoidea). The University of Kansas Science Bulletin, 39: EHRLICH, P. R The integumental anatomy of the silver-spotted skipper, Epargyreus clarus Cramer (Lepidoptera Hesperiidae). Microentomology, 24: GOMEZ, S Ciclo de desarrollo y hospederos de Heraclides anchisiades anchisiades (Lepidoptera: Papilionidae). Um modelo exploratório para evaluar la sostentabilidad de la cria de mariposas ornamentales em la comunidad indígena de Pena Roja. Revista Colombiana de Entomologia, 28 (11):
76 HALL, J. P. W A revision of the genus Theope its systematics and biology (Lepidoptera: Riondinidae: Nymphidiini). USA, Scientific Publishers, 127 p. HALL, J. P. W A revision of the new riodinid butterfly genus Dachetola (Lepidoptera: Riodinidae). Journal of the New York Entomological Society, 109 (2). HALL, J. P. W Phylogenetic reassessment of the five forewing radial-veined tribes of Riodininae (Lepidoptera: Riodinidae). Journal of the Royal Entomology Society, 28: HALL, J. P. W. & C. J. CALLAGHAN A revision of the new riodinid butterfly genus Pseudotinea (Lepidoptera: Riodinidae). Journal of Natural History, 37: HALL, J. P. W. & D. J. HARVEY Basal subtribes of the Nymphidiini (Lepidoptera: Riodinidae): phylogeny and myrmecophily. Cladistics 18 (4): HANCOCK, D. L Classification of the Papilionidae (Lepidoptera): A phylogenetic approach. Smithersia, 2: JOHNSON, K. & R. ROZYCKI A new specie of the anchisiades group of Heraclides from Venezuela (Lepidoptera: Papilionidae). Journal of the New York Entomological Society, 94 (3): , illustr. KLOTS, A. B Lepidoptera, pp In S. L. Tuxen [ed.], Taxonomists glossary of genitalia in insects. Munksgaard, Copenhagen, Denmark. 283 p. 65
77 KRISTENSEN, N. P Skeleton and Muscles: Adults, p In: P. N. KRISTENSEN (Ed.). Band/Volume IV Arthropoda: Insecta. Teilband / Part 36. Lepidoptera moths and butterflies: Morphology, Physiology and Development. Vol. 2, X+564p. In: M. FISCHER (Ed.). Handbook of Zoology. Berlin, New York, Walter de Gruyter. KRISTENSEN, N. P; M. J. SCOBLE & O. KARSHOLT Lepidoptera phylogeny and systematics: the state of inventorying moth and butterfly diversity. Zootaxa, 1668: KUZNETZOV, V. I. & A. A. STEKOL NIKOV Comparative Morphology of Male Genitalia in the Subfamilies Morphinae and Brassolinae and Resurrection of the Family Satyridae, status resurr. (Lepidoptera), in a New Sense. Entomological Review, 81 (4): LAMAS, G Atlas of Neotropical Lepidoptera. Checklist: part 4A. Hesperioidea Papilionoidea. Vo. 5A. USA. Scientific publishers, 439p. LOPES, L. de O; A. P. B. MOTA & A. L. M. MESQUITA Determinação dos períodos pré-pupal e pupal da lagarta dos citros Papilio anchisiades capys (Hübner, 1809). Fitossanidade, 3 (1-2): 49. MADDEN, A. H The external morphology of the Tobacco Hornworm (Lepidoptera, Sphingidae). Annals of the Entomological Society of America, 37: MATSUDA, R Morphology and evolution of the insect head. The American Entomological Institute, 4: MATSUDA, R Morphology and Evolution of the Insect Thorax. Memoirs of the Entomological Society of Canada, 76: MATSUDA, R Morphology and Evolution of the Insect Abdomen. Oxford, Pergamon Press, VIII + 534p. 66
78 MIELKE, C. G. C; O. H. H. MIELKE & M. M. CASAGRANDE Estudo comparado de morfologia externa de Zaretis itys itylus (Westwood) e Agrias claudina anneta (Gray) (Lepidoptera, Nymphalidae, Charaxinae). I. Cabeça, apêndices e região cervical. Revista Brasileira de Zoologia, 21 (2): MIELKE, C. G. C; O. H. H. MIELKE & M. M. CASAGRANDE Estudo comparado da morfologia externa de Zaretis itys itylus (Westwood) e Agrias claudina anneta (Gray) (Lepidoptera, Nymphalidae, Charaxinae). II. Tórax e apêndices. Revista Brasileira de Zoologia, 21 (3) MIELKE, C. G. C; O. H. H. MIELKE & M. M. CASAGRANDE Estudo comparado de morfologia externa de Zaretis itys itylus (Westwood) e Agrias claudina annetta (Gray). (Lepidoptera, Nymphalidae, Charaxinae) III. Abdome. Revista Brasileira de Zoologia, 21 (4): MILLER, James S Phylogenetic studies in the Papilioninae (Lepidoptera: Papilionidae). Bulletin of the American Museum of Natural History 186: MILLER, Jacqueline Y The head capsule of selected Hesperioidea. Journal of Research on the Lepidoptera. California, U.S.A. 9 (4): (1971). MUNROE, E. G The classification of the Papilionidae (Lepidoptera). The Canadian Entomologist. Supplement. 17: OITICICA, J. F Sobre a morfologia do pênis em Lepidoptera. Boletim do Museu Nacional, 50: OLIVEIRA, B. D. L. & D. URBAN Contribuição ao conhecimento da biologia de Papilio anchisiades capys (Hübner, 1809) (Lepidoptera Papilionidae). Dusenia, 10 (2): 89-95, 6 fig. 67
79 PENZ, C. M. & P. J. DEVRIES Preliminary Assessment of the Tribe Lemoniini (Lepidoptera Riodinidae) Based on Adult Morphology. American Museum Novitates, 3284: PENZ, C. M. & P. J. DEVRIES Systematic position of Apodemia paucipuncta (Riodinidae), and a critical evaluation of the Nymphidiine transtilla. Zootaxa, 1190:1-50. PIERCE, F. N The genitalia of the group Noctuidae of the Lepidoptera of the British Islands. Liverpool, Duncan. 88p. + 32pl. PIERCE, F. N The genitalia of the group Geometridae of the Lepidoptera of the British Islands. Faringdon, Oxon, 84p + 48pl. SCOBLE, M. J The Lepidoptera : form, function and diversity. Natural History Museum Publications. Oxford University Press, 404p. SHEPARD, H The pleural and sternal sclerites of the Lepidopterous thorax. Annals of Entomological Society of America, 23: SNODGRASS, R. E Principles of insect morphology. McGraw-Hill Book Company. New York and London. 667 pp., 319 figs. SORENSEN, J. T An integumental anatomy for the butterfly Glaucopsyche lygdamus (Lepidoptera: Lycaenidae): a morphological terminology and homology. Zoological Journal of the Linnean Society, 70: SRIVASTAVA, K. P Studies on the lemon butterfly Papilio demoleus L. (Lepidoptera). II Skeleto muscular mechanism (cervix and prothorax). Indian Journal of Entomology, 23:
80 SRIVASTAVA, K. P Studies on the lemon butterfly Papilio demoleus L. (Lepidoptera). III Skeleto muscular mechanism (pterothorax and its legs). Indian Journal of Entomology, 24: SRIVASTAVA, K. P Studies on the lemon butterfly Papilio demoleus L. (Lepidoptera). Part V. Skeleto muscular system of the abdomen. Zoologischer Anzeiger, 177: STEHR, F. W Order Lepidoptera, p In: F. W. STEHR (Ed.). Immature Insects. Duboque, Kendall Hunt, 754p. TYLER, H; K. S. BROWN & K. WILSON Swallowtail Butterflies of the Americas - A Study in Biological Dynamics, Ecological Diversity, Biosystematics, and Conservation. Scientific Publishers, Gainesville. 376 p. YOUNG, A. M; M. S. BLUM; M. M. FALES & Z. BRIAN Natural history and ecological chemistry of the Neotropical butterfly Papilio anchisiades (Papilionidae). Journal of the Lepidopterist s Society, 40 (1): 36-53, 7 fig. 69
81 CAPÍTULO II MORFOLOGIA, COMPORTAMENTO, PARASITISMO E MECANISMOS DE DEFESA DOS IMATUROS DE Heraclides anchisiades capys (Hübner, [1809]) (LEPIDOPTERA : PAPILIONIDAE) RESUMO A importância ambiental e ecológica dos lepidópteros é considerável pelo fato de suas larvas se alimentarem de plantas e porque todos os estágios desses insetos podem ser consumidos por predadores ou parasitóides. Heraclides anchisiades capys é comum dentro de Papilionidae, alimentando-se de Citrus spp. (Rutaceae). Possui grande potencial econômico e facilidade de criação em cativeiro. Contudo, até os dias atuais são raras as informações sobre sua biologia. O estudo tem como objetivo caracterizar morfologicamente os imaturos relacionando-os com os diferentes estágios de desenvolvimento, registrar aspectos do comportamento, parasitismo, e mecanismos de defesa. Exemplares foram trazidos de Rio Vermelho, São Bento do Sul, Santa Catarina, acondicionados em gaiolas de criação e casa de vegetação juntamente à planta hospedeira. Foram realizados desenhos morfológicos com auxílio de câmara clara, fotografias dos diferentes estágios e microscopia eletrônica de varredura de algumas estruturas. Dentre os resultados observou-se que os ovos possuem formato esférico, sendo recobertos por substância amareloalaranjada secretada pela fêmea. Foram registrados seis ínstares larvais, e em todos, gregarismo durante o dia e dispersas para se alimentarem durante a noite. São parasitados por Pedinopelte gravenstii (Guérin - Ménéville, 1826) (Hymenoptera : Ichneumonidae) que emerge das pupas através de um orifício circular. As pupas de aspecto longo e delgado apresentam três padrões distintos de coloração associados ao processo de camuflagem com a planta hospedeira. Duração mínima dos estágios: ovo: 16 dias, 1º ínstar: 5 dias, 2º ínstar: 4 dias, 3º ínstar: 5 dias, 4º ínstar: 5 dias, 5º ínstar: 4 dias, 6º ínstar: 8 dias e 2 dias em pré-pupa, período pupal: 24 dias. Palavras - chave: Bionomia; Ciclo de Vida; Papilioninae; Papilionoidea. 70
82 MORPHOLOGY, BEHAVIOUR, PARASITISM AND MECHANISMS OF DEFENSE OF THE IMMATURES OF Heraclides anchisiades capys (Hübner, [1809]) (LEPIDOPTERA : PAPILIONIDAE) ABSTRACT The environmental and ecological importance of Lepidoptera is considerable due to the fact that its caterpillars feed on plants and because all stages of theses insects can be consumed by predators or parasites. Heraclides anchisiades capys is common among the Papilionidae, feeding on Citrus spp. (Rutaceae). It has a relevant economical potential and it is easily raised in captivity. However, until now information about its biology is rare. The study focuses on characterizing morphologically the immatures, relating them with the different stages of the development, register behavioral aspects, parasitism, and mechanisms of defense. Samples were brought from Rio Vermelho, São Bento do Sul, Santa Catarina, kept in creation cages and greenhouse together with the host plant. Morphological drawings with the help of light chamber, photos of several stages and scanning electronic microscopy of some structures were done. Among the results, the eggs have spherical shape, being covered by a yellowish-orange substance secreted by the female. Six larval stages were registered, and in all, gregarism during the day and spred to eat during the night. Parasited by Pedinopelte gravenstii (Guérin - Ménéville, 1826) (Hymenoptera : Ichneumonidae) that emerges from pupae through a circular hole. Pupae are long and slender with three distinct patterns of coloring associated to the camouflage process with the host plant. Minimum duration of the stages: egg: 16 days, 1 st instar: 5 days, 2 nd instar: 4 days, 3 rd instar: 5 days, 4 th instar: 5 days, 5 th instar: 4 days, 6 th instar: 8 days and 2 days in pre-pupae, pupal stage: 24 days. Key words: Bionomy; Life cycle; Papilioninae; Papilionoidea. 71
83 1. Introdução De acordo com DIAS (2006), na grande maioria dos lepidópteros do Neotrópico os imaturos são desconhecidos ou então parcialmente descritos, com informações apenas sobre a larva de último ínstar, pupa e planta hospedeira. HASENFUSS & KRISTENSEN (2003) consideram os imaturos e seus aspectos morfológicos como instrumentos de importância crucial para o estabelecimento das relações filogenéticas. CASAGRANDE & MIELKE (2005) ressaltam a importância do conhecimento dos imaturos para estudos de taxonomia de Lepidoptera em seus níveis superiores e suas relações. Segundo STEHR (1987), a família Papilionidae possui imaturos relativamente bem conhecidos por conter espécies conspícuas, cujas larvas são confiavelmente associadas aos adultos, assim como nas espécies de Saturniidae, Sphingidae e Nymphalidae. Alguns poucos estudos abordaram aspectos referentes aos imaturos das espécies Neotropicais de Papilionidae (D ALMEIDA 1922; 1966; PENZ & ARAÚJO 1990; TYLER et al. 1994) e especificamente a Heraclides anchisiades capys (Hübner, [1809]) (OLIVEIRA & URBAN 1977; LOPES et al. 1979; BASTOS 1980; MOTA & BASTOS 1980) e para Heraclides anchisiades idaeus (Fabricius, 1793) YOUNG et al. (1986), portanto realmente escassos. Também são raros dentro de Papilionidae estudos que abordem detalhadamente aspectos morfológicos e comportamentais dos imaturos, e que vêm a ser dados de grande importância taxonômica, como os apresentados em outros artigos que abordam estes aspectos em outros Papilionoidea e Hesperioidea (COMSTOCK & VÁZQUEZ 1961; CASAGRANDE 1979; YOUNG 1985; SILVA et al. 2006; SOUZA et al. 2006; KAMINSKI et al. 2008). 72
84 2. Objetivos A carência de estudos mais aprofundados em relação aos estágios imaturos de Lepidoptera e a importância do conhecimento destas fases de vida para a sistemática dos lepidópteros tornam o presente estudo de grande valia. O propósito do trabalho foi caracterizar morfologicamente os imaturos e registrar aspectos do comportamento, parasitismo e mecanismos de defesa, buscando um enfoque comparativo com os demais Papilionoidea e Hesperioidea conhecidos. 73
85 3. Material e Métodos Os exemplares foram trazidos de Rio Natal, São Bento do Sul, Santa Catarina, logo após a postura, juntamente com a planta hospedeira Citrus sp., sendo então mantidos no laboratório em gaiola de criação, acomodados sobre papel absorvente, este último borrifado com água mineral diariamente para a manutenção da umidade até o momento da eclosão. As larvas foram mantidas em gaiolas de criação e observadas periodicamente, os galhos de Citrus sp. foram trocados diariamente para a alimentação dos indivíduos, assim como a limpeza das gaiolas. Na última idade as larvas foram transferidas para casa de vegetação e mantidas em plantas jovens até a formação das pupas e emergência dos adultos. Para cada fase do desenvolvimento foram preservados três indivíduos. As larvas foram sacrificadas em água fervente, fixadas em líquido KAHLE-DIETRICH por um período de três dias e então transferidos para álcool 70%. Cápsulas cefálicas obtidas a cada mudança de ínstar foram mantidas a seco em recipientes plásticos. Exemplares foram descritos e fotografados a cada novo evento, com o auxílio de câmera digital, técnicas de fotografia em auto-montagem e microscopia eletrônica de varredura. Os desenhos e a quetotaxia foram feitos através de microscópio estereoscópico acoplado à câmara clara. 3.1 Terminologia Utilizada Para as estruturas da larva e quetotaxia utilizou-se principalmente a terminologia utilizada por STEHR (1987), enquanto que para as pupas DIAS (2006). Em ambos os casos com modificações frente aos trabalhos clássicos e outros mais recentes que abordaram a morfologia dos imaturos (MOSHER 1916; PETERSON 1962; CASAGRANDE 1979; SCOBLE 1992; PAIM et al. 2004). 3.2 Considerações sobre a terminologia da larva O termo escudo cervical é adotado por alguns autores para a região do pronoto das larvas que se apresenta em forma de uma placa fortemente esclerotinizada (PETERSON 1962; MARCONATO & DIAS 2004; DIAS 2006). 74
86 STEHR (1987) e SCOBLE (1992) denominam a mesma de escudo protorácico. Placa protorácica é a terminologia seguida por alguns autores para a mesma estrutura (PARSONS 1996; PAIM et al. 2004; ZENKER et al. 2007). Adota-se o termo placa pronotal neste estudo, por considerar este o mais adequado para a supracitada região, seguindo alguns autores (OLIVEIRA & URBAN 1977; 1979; 1987; CASAGRANDE 1979). Vários autores denominam de larvópodos os apêndices locomotores pertencentes ao abdome (URBAN & OLIVEIRA 1972; 1980; SILVA et al. 2006; ZENKER et al. 2007). O termo falsas pernas é utilizado em SOUZA et al. (2006). Prolegs refere-se às mesmas estruturas, sendo utilizado em alguns trabalhos (PETERSON 1962; SCOBLE 1992; FREITAS 2006; KAMINSKI & FREITAS 2008). SPECHT et al. (2007) adota pseudópodos como o termo mais adequado. O trabalho de CASAGRANDE (1979) trata as estruturas citadas como pernas abdominais, sendo que FURTADO (2004) adota pernas abdominais para aquelas pertencentes aos segmentos 3 a 6 do abdome e pernas anais para aquelas localizadas no último segmento abdominal. Adota-se neste estudo pernas abdominais como o termo mais adequado, levando em consideração sua localização nesta parte do corpo. Placa anal é o termo utilizado no presente estudo referindo-se a estrutura dorsal, esclerotinizada do último segmento abdominal, segue grande parte dos autores (YOUNG 1985; SCOBLE 1992; LORINI & CORSEUIL 2001; ZENKER et al. 2007). STEHR (1987) preferiu denominar a região de escudo anal. Outros fazem uso do termo placa supra-anal (URBAN & OLIVEIRA 1989; FURTADO 2004), sendo que os trabalhos de MARCONATO & DIAS (2004) e DIAS (2006) se referem a esta estrutura como escudo supra-anal. Placa suranal também é adotada em alguns trabalhos de morfologia (PETERSON 1962; CASAGRANDE 1979; OLIVEIRA & URBAN 1987; CASAGRANDE & MIELKE 2005). 3.3 Considerações sobre a terminologia da pupa Os trabalhos de SPECHT et al. (2007) e ZENKER et al. (2007) fazem uso do termo podoteca e pteroteca para as pernas e asas respectivamente. No presente estudo adota-se a terminologia mais usual, denominando de perna e asas as estruturas citadas, o que segue a maioria dos autores (CASAGRANDE 1979; 75
87 SCOBLE 1992; FURTADO 2004; PAIM et al. 2004; CASAGRANDE & MIELKE 2005; SILVA et al. 2006). 76
88 4. Resultados 4.1 Aspectos morfológicos Ovo (Figs 1-7) Esférico, sem ornamentações e com pólo inferior plano em contato com o substrato. Recoberto por substância de coloração amarelo-alaranjada, secretada pela fêmea durante a oviposição e mais abundante na região mediana; lateralmente formando faixas mais espessas arqueadas, circundando parcialmente o ovo, da face inferior para a superior. Área micropilar arredondada e evidente na face superior. Diâmetro médio: 1,10 mm. Duração mínima: 16 dias Larva de 1º ínstar (Figs 8-15) Cabeça arredondada, castanho com muitas cerdas e sem escolos ou ornamentações. Frontalmente a sutura epicranial divide a cabeça em três regiões, os epicrânios lateralmente onde se encontra a maior parte das cerdas primárias, e medianamente a fronte, delimitada lateralmente pela sutura epicranial e inferiormente pelo clípeo, este em forma de banda esclerotinizada transversal, limitada ventralmente pelo anteclípeo membranoso. Ínfero-medianamente, o labro bilobado e ligado ao anteclípeo superiormente. Mandíbulas fortemente esclerotinizadas, subtriangulares e côncavas oralmente sem borda denteada. Seis estemas látero-inferiormente com tamanhos semelhantes, sendo 1, 2, 3, 4 e 6 dispostos em semicírculo, estema 5 isolado ventralmente e próximo à base da antena, esta com três artículos, sendo o apical reduzido e o mediano o maior dentre os três. Placa pronotal castanho-alaranjado, subretangular e com vinte pares de cerdas sobre calazas, o restante do protórax de coloração alaranjada. Pernas torácicas semelhantes e compactas, formadas por coxa, trocânter, fêmur, tíbia e tarso unisegmentado com garra terminal e com fortes cerdas dispostas irregularmente. 77
89 Tegumento do corpo pardo-oliva com tonalidades mais claras nas regiões inter-segmentais. Placa anal, subtriangular com a mesma coloração da placa pronotal e com dez pares de cerdas sobre calazas. Pernas abdominais de A3 A6 com ganchos dispostos em mesosérie uniordinal e laterosérie uniordinal, em A10 com ganchos em penelípse uniordinal. Mensurações: Cápsula cefálica: largura 0,8 mm; altura 0,7 mm. Comprimento médio da larva: 4,5 mm. Duração mínima: 4 dias. Quetotaxia da Cabeça (Figs 9-11) Vinte pares de cerdas primárias, excluindo aquelas pertencentes ao labro. Grupo Clipeal (C): C1 diminuta e próxima à linha mediana sagital da cabeça; C2 maior e próxima à margem ventral do clípeo. Grupo Frontal (F): F1, cerda única neste esclerito, próxima à sutura epicranial; poro Fa na mesma altura de F1 e próximo da linha média sagital. Grupo Adfrontal (Af): Af2 dorsal à Af1 na adfronte, ambas apresentando o mesmo comprimento; poro Afa não observado. Grupo Anterior (A): A1 dorsal à antena, situada medianamente entre o estema 4 e a adfronte; A2 dorsal à A1 na mesma altura de Af1; A3 dorsal à A2 e aos estemas 1 e 2; poro Aa látero-ventral à A2. Grupo Estemal (E): E1 lateral ao estema 3; E2 dorsal ao estema 1; E3 localizado lateralmente ao semicírculo estemal; poros Ea e Eb não observados. Grupo Subestemal (Se): Se1 ventral ao estema 5 e mais próxima do alvéolo antenal; Se2 látero-dorsal à Se1; Se3 látero-ventral à Se2. Todas as cerdas subestemais de comprimento semelhante. Grupo Lateral (L): L1 dorsal à E2; poro La não observado. Grupo Póstero-dorsal (P): P1 entre L1 e Af2 e mais próxima desta; P2 láterodorsal à P1 e de comprimento menor; poro Pa ventral à P1; poro Pb não observado. Grupo de Micro-cerdas dorsais (Md): Md1 dorsal à P2 e tão próxima desta como da sutura epicranial; Md2 dorsal à Md1 e mais próxima da sutura epicranial; poro Mda lateral e na mesma altura de Md2; Md3 dorsal ao poro Mda. 78
90 Quetotaxia do Tórax (Fig. 12) Protórax: Oito pares de cerdas. Xd1 e Xd2 dispostas na margem anterior da placa pronotal, Xd1 dorsal à Xd2; D1 e D2 na margem posterior da placa pronotal, D1 dorsal à D2. L1 e L2 dispostas em verruga ântero-ventralmente ao espiráculo, L1 dorsal à L2. Grupo supra-ventral (SV) em verruga dorsal à perna, SV1 dorsal à SV2. Meso e Metatórax: Oito pares de cerdas, todas dispostas sobre verrugas e sobre calazas, exceto D1, esta dorsal e anterior à linha média transversal, disposta sobre calaza, D2 maior e póstero-ventral à D1. Grupo subdorsal (SD) ventral à D2 e póstero-dorsal ao grupo Lateral (L), SD1 dorsal à SD2. L1 dorsal e pouco maior que L2. Grupo (SV) como no protórax. Quetotaxia do Abdome (Fig. 13) A1 A2: Doze pares de cerdas, estando D2, SD1, SD2, SV1 e SV2 localizadas sobre verrugas. D1 ântero-dorsal à D2. Grupo (SD) ântero-ventral à D2 e dorsal ao espiráculo, SD1 dorsal à SD2. Cerdas laterais póstero-ventrais ao espiráculo, sendo L1 maior e ântero-dorsal à L2. SV1 ventral ao espiráculo e SV2 póstero-ventral à SV1. MV1 posterior à SV2 e na mesma altura, MV2 póstero-ventral à SV2 e MV3 ântero-ventral à MV2. Cerda ventral V1 póstero-ventral à MV2. A3 A5: Onze pares de cerdas com distribuição semelhante às de A1 e A2, exceto os grupos (SV) em única verruga com SV1 dorsal à SV2 e o grupo microventral (MV) dorsal à perna, sendo MV1 póstero-dorsal à MV2 e esta última posterior à MV3. A6: Doze pares de cerdas. L3 anterior ao grupo supra-ventral (SV), ânteroventral em relação às outras cerdas laterais. As demais com distribuições semelhantes às de A3, A4 e A5. A7 A8: Doze pares de cerdas. V1 ventral à MV1 e o restante das cerdas com a mesma conformação de A3 A5. A9 + A10: Nove pares de cerdas. Na região de A9, D1 ântero-dorsal à D2, SD1 ventral à D2, L1 póstero-ventral à SD1 e SV1 póstero-ventral à L1. Na região de A10, D1 e D2 na margem dorsal da placa anal, sendo D1 maior e dorsal à D2; SD1 e SD2 próximas à margem ventral da placa anal com SD2 anterior à SD1, ambas de comprimentos semelhantes. 79
91 4.1.3 Larva de 2º ínstar (Figs 16 e 17) No início as larvas apresentam cor amarelada uniforme em toda extensão, com mudanças após vinte e quatro horas. Cabeça castanho-amarelado, com estreita borda enegrecida na margem posterior; região apical das mandíbulas escurecida. Placa pronotal amarelo-castanho e o restante do protórax amarelo-oliva, pernas torácicas com a mesma cor da placa pronotal. Meso, metatórax e A1 - A6, amarelo translúcidos com regiões inter-segmentais amarelo intenso; A7 até A10 com tonalidade amarelada. Pernas abdominais de A3 - A6 com os ganchos em mesosérie biordinal e laterosérie uniordinal, A10 com ganchos em penelípse biordinal. Mensurações: Cápsula cefálica: largura 1,2 mm; altura 1,06 mm. Comprimento médio da larva: 10 mm. Duração mínima: 5 dias Larva de 3º ínstar (Figs 18 e 19) Cabeça castanho-amarelado; placa pronotal e pernas protorácicas amarelocastanho. Tegumento do tórax e abdome com tonalidades entre amarelo e verdeoliva, sendo o amarelo mais intenso na região do protórax, áreas inter-segmentais e de A7 até A10. Pernas abdominais de A3 - A6 com ganchos agrupados em mesosérie triordinal e laterosérie biordinal, em A10 dispostos em penelípse triordinal. Mensurações: Cápsula cefálica: largura 1,73 mm; altura 1,6 mm. Comprimento médio da larva: 15,3 mm. Duração mínima: 5 dias Larva de 4º ínstar (Figs 20 e 21) Cabeça castanho; placa pronotal amarelo-esverdeada e pernas torácicas na mesma tonalidade da cabeça. Tegumento do tórax e abdome verde-oliva, passando ao amarelo-esverdeado a partir de A7. Região ventral do tórax e abdome brancoacinzentada, sendo esta mesma tonalidade observada lateralmente nas pernas abdominais de A3 - A6. Placa anal castanho-amarelado. Abdome com manchas brancas difusas distribuídas de A2 - A9, sendo duas faixas a partir do ápice de A2 80
92 formando desenhos irregulares; manchas dorsalmente dispostas em A3, A4, A8 e A9; lateralmente com manchas arredondadas dispostas acima e abaixo dos espiráculos de A2 - A6, conspícuas em A3 e em A2 sobrepostas à margem distal. Base das pernas abdominais com faixa branca arqueada. Escolos do protórax amarelo-esverdeados, os demais verde-oliva; base dos escolos abdominais com mancha esbranquiçada, exceto nos dois primeiros segmentos. Ganchos da planta das pernas abdominais semelhantes aos da terceira idade. Mensurações: Cápsula cefálica: largura 2,8 mm; altura 2,6 mm. Comprimento médio da larva: 29,4 mm. Duração mínima: 5 dias Larva de 5º ínstar (Figs 22, 23, 39, 40) Cabeça castanho com sutura epicranial branca, área dos estemas creme. Placa pronotal castanho-avermelhado com faixa branca medianamente, manchas de cor creme, laterais à faixa média e com escolos ântero-laterais castanhoamarelados. Tegumento do tórax e abdome castanho-esverdeado, sendo brancoacinzentado na região ventral, incluindo a planta das pernas abdominais. Placa anal castanho. Manchas brancas distribuídas desde o tórax até A9, formando losângulos ao longo do dorso, conspícuas a partir da margem distal de A2 até a base de A9. Manchas brancas arredondadas distribuídas lateralmente de forma irregular, sendo distintas em A3, A4, A7 - A9. Restante do tegumento dorsal e lateral marcado por finas faixas e pontos brancos. Escolos do corpo castanho-amarelados com base branca, exceto os protorácicos sem a base branca. Ganchos das pernas abdominais semelhantes aos ínstares terceiro e quarto. Mensurações: Cápsula cefálica: largura 3,2 mm; altura 3,0 mm. Comprimento médio da larva: 37,6 mm. Duração mínima: 4 dias Larva de 6º ínstar (Figs 24, 25, 41-59) Cabeça castanho com sutura epicranial branca; manchas creme na área dos estemas e pequenas manchas arredondadas creme no vértice, região esta mais amarelada em relação ao restante da cabeça. Cor do tegumento do tórax e abdome, 81
93 distribuição dos escolos e ganchos das pernas abdominais semelhantes ao ínstar anterior, sendo os escolos maiores e mais amarelados com cerdas pequenas, esbranquiçadas e de tamanhos semelhantes. Manchas brancas dorsomedianamente formando losângulos conspícuos da margem distal de A1 até a base de A9, mais evidentes em A3, A4, A8 e A9, diferindo do ínstar anterior. Mensurações: Cápsula cefálica: largura 4,3 mm; altura 4,1 mm. Comprimento médio da larva: 58,23 mm. Duração mínima: 10 dias, sendo 2 dias em pré-pupa. Quetotaxia (Figs 51 e 52) Protórax: Escolo localizado ântero-ventralmente na placa pronotal, com cerdas Xd1 e Xd2; cerdas do grupo dorsal (D1 e D2) na margem posterior da placa, D1 dorsal à D2, L1 dorsal à L2, ambas no escolo ântero-ventral ao primeiro espiráculo. Meso e Metatórax: D1 sobre escolo dorsal, anterior à linha mediana transversal. D2 no escolo póstero-ventral ao escolo de D1. Escolo ântero-ventral com as cerdas do grupo subdorsal, SD1 e SD2. A1 A2: D1 e D2 semelhantes ao metatórax, ausência de SD1 e SD2. A3 A8: Escolo médio-dorsal inconspícuo. Cerda D2 localizada no escolo póstero-dorsal à linha mediana transversal, este mais dorsal em A7 e A8. A9 + A10: D2 no escolo, semelhante aos segmentos abdominais anteriores. Em A10, D1 e D2 na margem dorsal da placa anal, D1 ântero-dorsal à D2; SD1 e SD2 próximas à margem ventral da placa, SD2 ântero-ventral à SD Pupa (Figs 28-38, 60-70) Adéctica obtecta, sucinta. Alongada diminuindo gradativamente em direção ao cremaster. Tegumento fino e densamente rugoso, com pequenas excrescências dispostas irregularmente e com projeções onduladas. Com três padrões distintos de coloração: castanho-escuro; pardo-acinzentada com manchas de contorno definido e esverdeada. Cabeça prolongada anteriormente. Em vista ventral, duas fendas superiores separando a fronte das áreas parietais; clípeo elevado; fóveas tentoriais anteriores laterais ao clípeo, visíveis em microscopia eletrônica de varredura; região das 82
94 mandíbulas arredondada, ínfero-laterais ao clípeo e mais altas que este; labro pequeno e hexagonal, inferior ao clípeo e entre as mandíbulas; gáleas tão longas quanto as tecas das asas posteriores; antenas dispostas entre as pernas e as asas anteriores, unidas na linha mediana ventral, com tubérculos rugosos na base; área inferior do olho composto com tubérculo rugoso. Pronoto com profundas depressões tanto medianas como laterais, ânterolateralmente com dois tubérculos e posteriormente com vários tubérculos irregulares. Espiráculo elíptico na margem látero-posterior. Mesonoto com depressão na base e tubérculos irregulares, grandes protuberâncias ântero-laterais localizadas na base da asa anterior e com projeção arredondada dirigida anteriormente em carena, formando ângulo reto com a depressão basal. Metanoto, o menor dentre os segmentos do tórax, marcado por protuberâncias arredondadas, duas pequenas na margem basal e próximas à linha mediana longitudinal, duas grandes e duas médias lateralmente dispostas no terço médio, posteriormente a estas se encontra o cinto de sustentação, atravessando o terço posterior. Em vista ventral, a perna protorácica, lateral à gálea, com quatro tubérculos, um basal, um apical e dois medianos; perna mesotorácica com um tubérculo basal, um mediano e um apical. Asas anteriores com uma série de protuberâncias na margem externa e uma fenda que prende o cinto de sustentação no seu terço médio. Asas posteriores evidentes desde o metanoto, unindo-se medianoventralmente sob o ápice das gáleas. Abdome com dez segmentos, apresentando lateralmente espiráculos elípticos com borda interna serreada, visíveis de A2 - A8, sendo o par de A8 o menor. Dorsalmente A1 com quatro protuberâncias, duas na metade posterior próximas da linha mediana longitudinal e outras duas próximas às margens laterais. A2 com cinco protuberâncias, três na margem anterior, sendo uma mediana e duas laterais, as demais na metade posterior, próximas à linha média. A3 com a mesma conformação de A2, exceto pela ausência de uma protuberância ântero-mediana. A4 - A8 com número e distribuição de protuberâncias semelhantes à A1, sendo A4 - A6 os maiores dentre todos os segmentos abdominais e com as maiores protuberâncias. A9, o menor dos segmentos em vista dorsal, e juntamente com A10 apresentam pequenos tubérculos irregulares. Ventralmente é possível identificar caracteres de diferenciação entre os sexos, sendo as fêmeas possuidoras de sulco mediano longitudinal nos esternos A8 e A9 e os machos sem esta característica. 83
95 Cremaster posterior no A10 dividido em dois lobos achatados, os ganchos possuem coloração castanho-escuro e extremidade bífida, em forma de garra. Comprimento médio da pupa em ambos os sexos: 33,7 mm Duração mínima: 24 dias. 4.2 Comportamento, mecanismos de defesa e parasitismo. Uma vez fecundada, a fêmea procura a planta hospedeira Citrus sp. e permanece circundando a mesma de forma agitada por cerca de trinta minutos. Quando escolhido o local da postura, a fêmea pousa na superfície adaxial da folha e inicia o processo de oviposição, as asas anteriores estando inclinadas para trás, recurva o abdome em direção à superfície abaxial da folha, depositando um ovo e em seguida voltando o abdome para a posição normal; este procedimento é repetido até que toda postura seja efetuada. Nas posturas observadas o número de ovos variou de 71 a 93, levando cerca de trinta minutos para completar todo o processo. Os ovos são depositados lado a lado na folha, envoltos por uma substância amarelo-alaranjada que a fêmea secreta durante a oviposição e que recobre praticamente todo o cório. Larvas de 1º ínstar ao eclodirem alimentam-se do cório e desta substância amarelo-alaranjada de seus próprios ovos e daqueles que ainda não eclodiram, desta forma, os indivíduos que eclodem mais tarde se alimentam apenas do cório. Ao final do processo de eclosão a área de postura fica totalmente limpa, exceto por alguns ovos inviáveis que permanecem na folha. Larvas de 1º ínstar passam as primeiras horas agrupadas no centro da folha e posteriormente apresentam comportamento mais disperso, ora formando agregados em partes distintas da folha e sem regularidade em relação ao local, ora formando um único grupo, que passa a ser constante até o final do 4º ínstar. A maioria dos indivíduos permanece agregada durante o dia e dispersa durante a noite para se alimentar, retornando para as mesmas folhas onde tecem fios de seda, permanecendo reunidas. Alguns indivíduos se alimentam ocasionalmente durante o dia. A partir do 5º ínstar passam a se reunir no tronco principal da planta durante o dia e no restante do tempo com o mesmo comportamento dos ínstares anteriores. Antes da mudança de ínstar, os indivíduos se reúnem e permanecem totalmente imóveis, após a muda ingerem as exúvias, deixando apenas a cápsula cefálica. Nos diferentes ínstares, a rotina de alimentação ocorre sempre a partir das 84
96 margens da folha em movimentos semicirculares, consumindo toda a área foliar restando a nervura principal. O osmetério, glândula localizada no pronoto, com função de defesa contra os predadores, permanece retraída no pronoto, sendo protraída em qualquer ínstar quando se sentem ameaçadas. No primeiro ínstar conseguem protrair o osmetério, entretanto o odor exalado é pouco perceptível, o que se intensifica a partir do 2º ínstar. Esta estrutura, em todos os ínstares de coloração amarelada, com uma leve tonalidade alaranjada que se acentua na medida em que os ínstares vão se sucedendo. A superfície da glândula apresenta conformação rugosa com pequenas projeções delgadas distribuídas de forma irregular. No último ínstar, os indivíduos escolhem um local apropriado para darem início ao processo de empupamento, mas antes eliminam grande quantidade de bolos fecais. O local escolhido pode ser a própria planta hospedeira, outra planta vizinha ou até mesmo quaisquer substratos próximos ao local como paredes e muros (Fig. 27). A larva tece uma base de seda através de movimentos circulares para a direita e esquerda por cerca de vinte minutos, depois se volta prendendo os ganchos das pernas abdominais do último segmento na base de seda antes elaborada e então inicia o processo de construção do cinto de sustentação, primeiramente trabalhando nas bases para este cinto em ambos os lados do substrato e depois no próprio fio, reforçando-o várias vezes de uma base à outra, o que pode levar até uma hora. Em seguida, a larva passa a cabeça e o tórax por baixo do cinto e permanece fortemente curvada em pré-pupa por aproximadamente dois dias. Para a formação da pupa o corpo é afastado do substrato, e o tegumento rompido ao longo da linha média dorsal que percorre o pronoto e mesonoto, quando passa a se movimentar até livrar-se da exúvia larval por completo. As pupas permanecem imóveis e presas ao substrato até o momento da emergência do adulto. Três tipos de coloração foram observados, todos com estreita ligação ao padrão do substrato, assim os indivíduos que empupam na planta hospedeira são de difícil visualização por permanecerem completamente camuflados em meio aos galhos, folhas, musgos e liquens presentes, o que certamente lhes confere defesa contra possíveis predadores. Pupas com tegumento castanho-escuro se assemelham aos galhos secos ou quebrados, enquanto que as de coloração pardo- 85
97 acinzentada se confundem com os liquens do substrato e por fim, as de aspecto esverdeado, que se mesclam com os musgos presentes nos galhos, troncos e com as folhas. Após um período mínimo de vinte e dois dias, as pupas passam a uma coloração escurecida com o tegumento menos endurecido e, aproximadamente dois dias após este escurecimento, o tegumento se rompe na região dorsal da cabeça, por onde o adulto emerge com movimentos lentos, esticando o abdome ao mesmo tempo em que expele o mecônio, líquido denso e esbranquiçado, até se livrar completamente da exúvia pupal, quando então se desloca, caminhando até o ponto mais alto do galho e cadenciadamente inicia o esticamento das asas o que pode levar até dez minutos. Na maioria das vezes, fêmeas emergem antes que machos. Parasitismo foi constatado por Pedinopelte gravenstii (Guérin-Ménéville, 1826) (Hymenoptera, Ichneumonidae, Ichneumoninae). Um parasitóide emerge de cada pupa através de orifício circular ventro-lateralmente na altura das asas anteriores. 86
98 5. Discussão Estudos morfológicos e de história natural são de grande importância para a taxonomia dos lepidópteros, a necessidade do correto reconhecimento da espécie em qualquer estágio do ciclo de vida é de grande valia quando a mesma atinge o status de praga de alguma cultura, neste caso, a citricultura. O Brasil é um dos maiores produtores de citros e lidera o mercado mundial de suco de laranja. A laranja é a fruta mais produzida no território nacional e ocupa uma área plantada superior a 800 mil hectares (SANCHES et al. 2008). Citricultura é significantemente importante para a economia brasileira, desde que a mesma representa um ganho aproximado de 1.5 bilhões de dólares anualmente (PARRA et al. 2004). Devido à diversidade de cultivares e clima nos quais eles crescem, Citrus é provavelmente a árvore frutífera mais amplamente distribuída desde climas tropicais e subtropicais até habitats temperados pelo mundo. Conseqüentemente uma ampla diversidade de artrópodes pragas é registrada para as variedades de Citrus (SMITH & PEÑA 2002). Entre as potenciais pragas de Citrus está Heraclides anchisiades capys que quando ataca os brotos terminais causa grande estresse à planta com conseqüente comprometimento da qualidade dos frutos. O reconhecimento dos diferentes ínstares, aspectos bionomicos e morfológicos são fundamentais para suportar a adequada identificação da espécie, proposta deste trabalho. As características relacionadas aos ovos como: coloração, formato, disposição da substância que recobre o cório e duração do estágio, são significativos, visto que são exclusivas quando comparados com outras espécies de Papilionidae já estudados (COMSTOCK & VÁZQUEZ 1961; TYLER et al. 1994; PARSONS 1996; XIUSHAN et al. 2006) e entre os demais Papilionoidea e Hesperioidea documentados. Por exemplo, o formato, tamanho e duração do estágio de ovo, são caracteres fundamentais que os diferem de Heraclides anchisiades idaeus como observado por YOUNG et al. (1986), embora pertençam à mesma espécie. Hábitos observados nas larvas de 1º ínstar, como a primeira alimentação com a substância amarelo-alaranjada que recobre o cório dos ovos, assim como o uso do cório como primeiro alimento podem estar relacionados com o futuro desempenho, nos demais ínstares, ou até mesmo com o número maior de ínstares, no caso seis, 87
99 pouco usual em outros Papilionoidea, indicando que estas questões merecem estudos mais aprofundados. A morfologia dos ínstares larvais e em destaque a do 1º ínstar formam um conjunto de caracteres de importância taxonômica com particularidades únicas quanto à coloração, morfologia e quetotaxia da cabeça, tórax e abdome, principalmente em se tratando da posição das cerdas e da conformação dos ganchos das pernas abdominais, diferenciando-se dos demais Papilionoidea Neotropicais que têm seus ínstares larvais documentados morfologicamente (LAWRENCE & DOWNEY 1966; CASAGRANDE 1979; PAIM et al. 2004; SILVA et al. 2006; SOUZA et al. 2006; KAMINSKI et al. 2008). Em se tratando de estudos com a mesma espécie, a largura da cápsula cefálica e o número de ínstares larvais são caracteres variáveis. A largura das cápsulas cefálicas nos diferentes ínstares corrobora o estudo de MOTA & BASTOS (1980), entretanto o mesmo autor encontrou cinco ínstares larvais. OLIVEIRA & URBAN (1977) observaram seis ínstares larvais, o que é confirmado no presente trabalho, mas apresentando cápsulas cefálicas de largura superior. A presença de seis ínstares larvais também constatados por OLIVEIRA & URBAN (1977) é uma característica que difere os indivíduos deste estudo de Heraclides anchisiades idaeus publicado por YOUNG et al. (1986) e dos demais Papilionoidea e Hesperioidea por nós vistos. Parasitismo por Pedinopelte gravenstii (Hymenoptera Ichneumonidae) constatado por alguns autores (COSTA LIMA 1935; SILVA 1935) foi observado no presente estudo, sendo este mesmo parasitóide ainda não registrado para outros Papilionoidea. Os padrões de coloração observados nas pupas e que se relacionam diretamente com o substrato no qual estas se formam, corroboram estudos com este tipo de comportamento dentro da família Papilionidae (YOUNG et al. 1986; HAZEL & WEST 1996; STEFANESCU 2004; HIRAGA 2006). De acordo com JONES et al. (2007) esta capacidade de polimorfismo é única para os Papilioninae e ausente nos demais Papilionidae, entretanto encontrado em alguns Nymphalidae e Pieridae e sem registros para Hesperioidea. 88






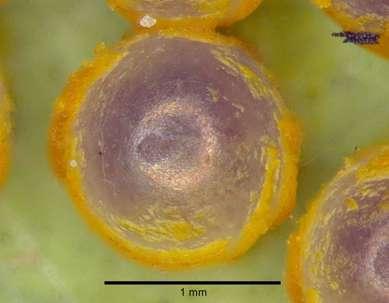
100 Figs 1-7. Heraclides anchisiades capys - Ovo: Imagem da postura vista lateral. 6 - Ovo em vista superior, com a substância amarelo-alaranjada. 7 - detalhe de um ovo com parte do cório aparente. 89
101 8 Fig. 8. Heraclides anchisiades capys: Larva de 1º ínstar em vista lateral. Lbr - labro. Md. - Mandíbula. Mx. - Maxila. 90

102 Figs Heraclides anchisiades capys: Larva 1º ínstar. 9, 11 - Cápsula cefálica em vista frontal Cápsula cefálica em vista lateral. An - antena. Md - mandíbula. Mx - maxila. Lbr - labro. 91
103 Esp. 12 Esp. Esp. Esp. Esp. 13 Figs 12 e 13. Heraclides anchisiades capys: Larva 1º ínstar Quetotaxia do tórax Quetotaxia do abdome. Esp - Espiráculo. 92






104 Figs Heraclides anchisiades capys: Larva. 14 e 15-1º ínstar. 16 e 17-2º ínstar. 18 e 19-3º ínstar. 93






105 Figs Heraclides anchisiades capys: Larva. 20 e 21-4º ínstar. 22 e 23-5º ínstar. 24 e 25-6º ínstar. 94





106 Figs Heraclides anchisiades capys Pré-pupa na planta hospedeira Pré-pupa em concreto Pupas: 28 - Padrão esverdeado Padrão castanho-escuro Padrão pardo acinzentado. 95








107 Figs Heraclides anchisiades capys. Pupas. 96






108 Figs Heraclides anchisiades capys. Larva: 39 e 40-5º ínstar: 39 - Estemas Detalhe de um estema º ínstar: 41 - Estruturas da cápsula cefálica em vista ventral Detalhe da região inferior da cápsula cefálica Detalhe da fiandeira e maxila Detalhe dos artículos antenais. An - antena. An1 - primeiro artículo antenal. An2 - segundo artículo antenal. An3 - terceiro artículo antenal. Clp - clípeo. Fi - fiandeira. Md - mandíbula. Mx - maxila. Lbr - labro. 97






109 Figs Heraclides anchisiades capys. Larva de 6º ínstar: 45 e 46 - Perna protorácica Perna abdominal do segmento A e 49 - Pernas abdominais dos segmentos A3 - A Detalhe dos ganchos da perna abdominal. Cx - coxa. Tr - trocânter. F - fêmur. Ti - tíbia. Ta - tarso. G - garra terminal. 98
110 Xd1 Xd2 SD1 SD2 Esp. 51 Esp. Esp. Esp. 52 Figs 51 e 52. Heraclides anchisiades capys: Larva 6º ínstar Quetotaxia do tórax Quetotaxia do abdome. Esp - Espiráculo. 99







111 Figs Heraclides anchisiades capys: Larva de 6º ínstar Cabeça e osmetério Osmetério Espiráculo abdominal. 100




.")

112 Figs Heraclides anchisiades capys: Pupa Superfície ventral da cabeça Detalhe lateral do olho e antena Espiráculo abdominal Porção distal da pupa vista ventral (A9, A10 e cremaster). 64 e 65 - Ganchos do cremaster. Clp - clípeo. Lbr - labro. Md - mandíbula. Fr - fronte. Ga - gálea. An - antena. O - olho. 101


113 66 67 Figs 66 e 67. Heraclides anchisiades capys: Pupa Macho esternos oito à dez Fêmea esterno oito à dez, destaque do sulco mediano. 102
114 68 69 Figs 68 e 69. Heraclides anchisiades capys. Pupa: 68 - Vista dorsal Vista ventral. 103
115 70 Fig. 70. Heraclides anchisiades capys. Pupa em vista lateral. 104

 (Hymenoptera, Ichneumonidae,")
116 71 72 Figs 71 e 72. Heraclides anchisiades capys. Exúvia pupal com orifício para saída do parasitóide Pedinopelte gravenstii (Guérin - Ménéville, 1826) (Hymenoptera, Ichneumonidae, Ichneumoninae). 105
GUSTAVO CAVALHEIRO AMARILIA
 GUSTAVO CAVALHEIRO AMARILIA MORFOLOGIA DO ADULTO E DOS IMATUROS DE Riodina lycisca (HEWITSON, [1853]), E SUAS RELAÇÕES COM A PLANTA HOSPEDEIRA (LEPIDOPTERA, RIODINIDAE). Tese apresentada à Coordenação
GUSTAVO CAVALHEIRO AMARILIA MORFOLOGIA DO ADULTO E DOS IMATUROS DE Riodina lycisca (HEWITSON, [1853]), E SUAS RELAÇÕES COM A PLANTA HOSPEDEIRA (LEPIDOPTERA, RIODINIDAE). Tese apresentada à Coordenação
Fernando M. S. Dias 1, Mirna M. Casagrande 1 & Olaf H. H. Mielke 1
 376 Morfologia do exoesqueleto de adultos de Memphis moruus stheno (Pritwittz) (Lepidoptera, Nymphalidae, Charaxinae) Fernando M. S. Dias 1, Mirna M. Casagrande 1 & Olaf H. H. Mielke 1 1 Departamento de
376 Morfologia do exoesqueleto de adultos de Memphis moruus stheno (Pritwittz) (Lepidoptera, Nymphalidae, Charaxinae) Fernando M. S. Dias 1, Mirna M. Casagrande 1 & Olaf H. H. Mielke 1 1 Departamento de
Luis Anderson Ribeiro Leite 1, Mirna Martins Casagrande 1 & Olaf Hermann Hendrik Mielke 1
 Morfologia, comportamento, parasitismo e mecanismos de defesa dos imaturos de Heraclides anchisiades capys (Hübner) (Lepidoptera, Papilionidae) 277 Luis Anderson Ribeiro Leite 1, Mirna Martins Casagrande
Morfologia, comportamento, parasitismo e mecanismos de defesa dos imaturos de Heraclides anchisiades capys (Hübner) (Lepidoptera, Papilionidae) 277 Luis Anderson Ribeiro Leite 1, Mirna Martins Casagrande
VINALTO GRAF ALICE FUMI KUMAGAI 3
 Acta Biol. Par., Curitiba, 33 (1, 2, 3, 4): 21-25. 2004 21 Uma espécie nova de Epirhyssa Cresson (Ichneumonidae, Hymenoptera) do Brasil 1 A new species of Epirhyssa Cresson, 1865 (Ichneumonidae, Hymenoptera)
Acta Biol. Par., Curitiba, 33 (1, 2, 3, 4): 21-25. 2004 21 Uma espécie nova de Epirhyssa Cresson (Ichneumonidae, Hymenoptera) do Brasil 1 A new species of Epirhyssa Cresson, 1865 (Ichneumonidae, Hymenoptera)
Larva de quinto estádio e pupa de Caligo martia ~Godart) (Lepidoptera, Nymphalidae, Brassolinae)
 (Lepidoptera, Nymphalidae, Brassolinae)") Revta bras. Zool. 17 (1): 75-79,2000 Larva de quinto estádio e pupa de Caligo martia ~Godart) (Lepidoptera, Nymphalidae, Brassolinae) Mirna M. Casagrande 2 Olaf H.H. Mielke 2 ABSTRACT. Fifth larval instar
Revta bras. Zool. 17 (1): 75-79,2000 Larva de quinto estádio e pupa de Caligo martia ~Godart) (Lepidoptera, Nymphalidae, Brassolinae) Mirna M. Casagrande 2 Olaf H.H. Mielke 2 ABSTRACT. Fifth larval instar
Estágios imaturos de Titaea orsinome Huebner (Lepidoptera, Saturniidae, Arsenurinae)
") Estágios imaturos de Titaea orsinome Huebner (Lepidoptera, Saturniidae, Arsenurinae) Eurides Furtado 1 ABSTRACT. Immature stagcs of Titaea orsinome Huebncr (Lepidoptera, Saturniidae, Arsenurinae). Data
Estágios imaturos de Titaea orsinome Huebner (Lepidoptera, Saturniidae, Arsenurinae) Eurides Furtado 1 ABSTRACT. Immature stagcs of Titaea orsinome Huebncr (Lepidoptera, Saturniidae, Arsenurinae). Data
Morfologia Externa Tórax e Abdome
 Morfologia Externa Tórax e Abdome Lucas Silva de Faria Morfologia Externa Revisão: Quais são os apêndices cefálicos??? Quais os tipos de aparelho bucal??? 1 Morfologia Externa Tórax 3 segmentos: Protórax
Morfologia Externa Tórax e Abdome Lucas Silva de Faria Morfologia Externa Revisão: Quais são os apêndices cefálicos??? Quais os tipos de aparelho bucal??? 1 Morfologia Externa Tórax 3 segmentos: Protórax
Nymphalidae, Ithomiinae) 1
 1") Morfologia ologia externa de Thyridia psidii cetoides (Rosenberg & Talbot). I. Cabeça e apêndices (Lepidoptera, Nymphalidae, Ithomiinae) 1 Jorge Manuel Saraiva Bizarro 2, Mirna Martins Casagrande 2 & Olaf
Morfologia ologia externa de Thyridia psidii cetoides (Rosenberg & Talbot). I. Cabeça e apêndices (Lepidoptera, Nymphalidae, Ithomiinae) 1 Jorge Manuel Saraiva Bizarro 2, Mirna Martins Casagrande 2 & Olaf
Estágios imaturos e bionomia de Cyclomia mopsaria Guenée (Lepidoptera, Geometridae) 1
 1") Estágios imaturos e bionomia de Cyclomia mopsaria Guenée (Lepidoptera, Geometridae) 1 Estágios imaturos e bionomia de Cyclomia mopsaria Guenée 1 Gláucia Marconato 2 & Manoel Martins Dias 3 1 Realizado
Estágios imaturos e bionomia de Cyclomia mopsaria Guenée (Lepidoptera, Geometridae) 1 Estágios imaturos e bionomia de Cyclomia mopsaria Guenée 1 Gláucia Marconato 2 & Manoel Martins Dias 3 1 Realizado
Ordem Lepidoptera. Ordem Lepidoptera. Ordem Lepidoptera. Borboletas e mariposas
 Borboletas e mariposas Asas membranosas cobertas por escamas que se destacam facilmente Aparelho bucal sugador maxilar que fica enrolado em repouso (espirotromba) As formas jovens (larvas) são denominadas
Borboletas e mariposas Asas membranosas cobertas por escamas que se destacam facilmente Aparelho bucal sugador maxilar que fica enrolado em repouso (espirotromba) As formas jovens (larvas) são denominadas
Morfologia externa comparada das três espécies do complexo Telchin licus (Drury) (Lepidoptera, Castniidae) com uma sinonímia
 (Lepidoptera, Castniidae) com uma sinonímia") (Drury) (Lepidoptera, Castniidae) com uma sinonímia Morfologia externa comparada das três espécies do complexo Telchin licus 245 Simeão S. Moraes 1,2 & Marcelo Duarte 2 1 Curso de Pós-Graduação em Ciências
(Drury) (Lepidoptera, Castniidae) com uma sinonímia Morfologia externa comparada das três espécies do complexo Telchin licus 245 Simeão S. Moraes 1,2 & Marcelo Duarte 2 1 Curso de Pós-Graduação em Ciências
Dardarina angeloi sp. n. do sudeste e sul do Brasil (Lepidoptera: Hesperiidae) 1
 1") Lundiana 6(supplement):31-35, 2005 2005 Instituto de Ciências Biológicas - UFMG ISSN 1676-6180 Dardarina angeloi sp. n. do sudeste e sul do Brasil (Lepidoptera: Hesperiidae) 1 Olaf H. H. Mielke & Mirna
Lundiana 6(supplement):31-35, 2005 2005 Instituto de Ciências Biológicas - UFMG ISSN 1676-6180 Dardarina angeloi sp. n. do sudeste e sul do Brasil (Lepidoptera: Hesperiidae) 1 Olaf H. H. Mielke & Mirna
Marina Moraes C. 1, Luis Anderson Ribeiro Leite 1, Mirna Martins Casagrande 1 & Olaf Hermann Hendrik Mielke 1
 Imaturos de Sarsina violascens (Herrich-Schäffer) (Lepidoptera, Noctuidae, Lymantriinae) 571 Marina Moraes C. 1, Luis Anderson Ribeiro Leite 1, Mirna Martins Casagrande 1 & Olaf Hermann Hendrik Mielke
Imaturos de Sarsina violascens (Herrich-Schäffer) (Lepidoptera, Noctuidae, Lymantriinae) 571 Marina Moraes C. 1, Luis Anderson Ribeiro Leite 1, Mirna Martins Casagrande 1 & Olaf Hermann Hendrik Mielke
Acta Biol. Par., Curitiba, 35 (1-2):
:") Acta Biol. Par., Curitiba, 35 (1-2): 63-67. 2006. 63 Uma espécie nova de Bicolletes Friese do nordeste brasileiro (Hymenoptera, Colletidae) 1 A new species of Bicolletes Friese fom Brazilian northeastern
Acta Biol. Par., Curitiba, 35 (1-2): 63-67. 2006. 63 Uma espécie nova de Bicolletes Friese do nordeste brasileiro (Hymenoptera, Colletidae) 1 A new species of Bicolletes Friese fom Brazilian northeastern
PRINCIPAIS ORDENS DE INSETOS
 PRINCIPAIS ORDENS DE INSETOS ORDENS DOS INSETOS ARCHAEOGNATHA traças saltadeiras. THYSANURA traça dos livros. ORDEM ORTHOPTERA Ortho = reta Ptera = asas Ordem que inclui gafanhotos grilos esperanças paquinhas
PRINCIPAIS ORDENS DE INSETOS ORDENS DOS INSETOS ARCHAEOGNATHA traças saltadeiras. THYSANURA traça dos livros. ORDEM ORTHOPTERA Ortho = reta Ptera = asas Ordem que inclui gafanhotos grilos esperanças paquinhas
GUATUBESIA, NOVO GÊNERO DE GONYLEPTIDAE
 Rev. bras. Ent. 22í2): 99-104 11.1X.1978 OPERA OPILIOLOGICA VARIA. XV. GUATUBESIA, NOVO GÊNERO DE GONYLEPTIDAE Helia E.M. Soares Guatubesia clarae, n. gen., n. sp. (Opiliones, Gonyleptidae, Gonyleptinae),
Rev. bras. Ent. 22í2): 99-104 11.1X.1978 OPERA OPILIOLOGICA VARIA. XV. GUATUBESIA, NOVO GÊNERO DE GONYLEPTIDAE Helia E.M. Soares Guatubesia clarae, n. gen., n. sp. (Opiliones, Gonyleptidae, Gonyleptinae),
8/23/2017. Apêndices locomotores: FACULDADE EDUCACIONAL DE MEDIANEIRA. Principais funções do tórax dos insetos: Pernas. Asas MORFOLOGIA DOS INSETOS
 FACULDADE EDUCACIONAL DE MEDIANEIRA MORFOLOGIA DOS INSETOS Profª. Dra. Patrícia Bellon. Agosto/2017 Principais funções do tórax dos insetos: Abriga os apêndices locomotores, pernas e asas. TÓRAX DO INSETO
FACULDADE EDUCACIONAL DE MEDIANEIRA MORFOLOGIA DOS INSETOS Profª. Dra. Patrícia Bellon. Agosto/2017 Principais funções do tórax dos insetos: Abriga os apêndices locomotores, pernas e asas. TÓRAX DO INSETO
Adeloneivaia schubarti Rêgo Barros & Mielke e seus estágios imaturos (Lepidoptera, Saturniidae, Ceratocampinae)
") Adeloneivaia schubarti Rêgo Barros & Mielke e seus estágios imaturos (Lepidoptera, Saturniidae, Ceratocampinae) Eurides Furtado 1 ABSTRACT. Adelolleivaia schubarti Rêgo Barros & Mielke and its immature
Adeloneivaia schubarti Rêgo Barros & Mielke e seus estágios imaturos (Lepidoptera, Saturniidae, Ceratocampinae) Eurides Furtado 1 ABSTRACT. Adelolleivaia schubarti Rêgo Barros & Mielke and its immature
Prepona pheridamas pheridamas (Cramer) e seus estágios imaturos (Lepidoptera, Nymphalidae, Charaxinae)
 e seus estágios imaturos (Lepidoptera, Nymphalidae, Charaxinae)") Prepona pheridamas pheridamas (Cramer) e seus estágios imaturos (Lepidoptera, Nymphalidae, Charaxinae) Eurides Furtado 1 ABSTRACT. Prepollll pheridllllllls pheridllllllls (Cramer) and its immalure stages
Prepona pheridamas pheridamas (Cramer) e seus estágios imaturos (Lepidoptera, Nymphalidae, Charaxinae) Eurides Furtado 1 ABSTRACT. Prepollll pheridllllllls pheridllllllls (Cramer) and its immalure stages
Aspectos Morfológicos de Lonomia obliqua Walker (Lepidoptera: Saturniidae) 1
 1") September, 2001 373 SYSTEMATICS, MORPHOLOGY AND PHYSIOLOGY Aspectos Morfológicos de Lonomia obliqua Walker (Lepidoptera: Saturniidae) 1 LISETE M. LORINI 2 E ÉLIO CORSEUIL 3 1 Parte da dissertação de Mestrado
September, 2001 373 SYSTEMATICS, MORPHOLOGY AND PHYSIOLOGY Aspectos Morfológicos de Lonomia obliqua Walker (Lepidoptera: Saturniidae) 1 LISETE M. LORINI 2 E ÉLIO CORSEUIL 3 1 Parte da dissertação de Mestrado
MORFOLOGIA EXTERNA TEGUMENTO - CABEÇA. Eng. Agr. Luiz Paulo
 MORFOLOGIA EXTERNA TEGUMENTO - CABEÇA Eng. Agr. Luiz Paulo Estrutura Geral do Tegumento É constituído de: membrana basal, epiderme e cutícula Tegumento - Origem Possui origem ectodérmica podendo ser descrito
MORFOLOGIA EXTERNA TEGUMENTO - CABEÇA Eng. Agr. Luiz Paulo Estrutura Geral do Tegumento É constituído de: membrana basal, epiderme e cutícula Tegumento - Origem Possui origem ectodérmica podendo ser descrito
Três espécies novas do gênero Chorisoneura (Blattellidae, Chorisoneuriinae) coletadas em ninhos de Sphecidae (Hymenoptera) do Estado do Acre, Brasil
 coletadas em ninhos de Sphecidae (Hymenoptera) do Estado do Acre, Brasil") Três espécies novas do gênero Chorisoneura (Blattellidae, Chorisoneuriinae)... 375 Três espécies novas do gênero Chorisoneura (Blattellidae, Chorisoneuriinae) coletadas em ninhos de Sphecidae (Hymenoptera)
Três espécies novas do gênero Chorisoneura (Blattellidae, Chorisoneuriinae)... 375 Três espécies novas do gênero Chorisoneura (Blattellidae, Chorisoneuriinae) coletadas em ninhos de Sphecidae (Hymenoptera)
Descrição dos Estágios Imaturos de Phthia picta (Drury) (Hemiptera: Coreidae)
 (Hemiptera: Coreidae)") June, 2001 Neotropical Entomology 30(2) 253 SISTEMÁTICA, MORFOLOGIA E FISIOLOGIA Descrição dos Estágios Imaturos de Phthia picta (Drury) (Hemiptera: Coreidae) RICARDO A. DA SILVA 1, PATRÍCIA S. FLORES
June, 2001 Neotropical Entomology 30(2) 253 SISTEMÁTICA, MORFOLOGIA E FISIOLOGIA Descrição dos Estágios Imaturos de Phthia picta (Drury) (Hemiptera: Coreidae) RICARDO A. DA SILVA 1, PATRÍCIA S. FLORES
FILO ARTHROPODA - INTRODUÇÃO
 Entomologia Entomologia veterinária: é o estudo de insetos de importância veterinária, num sentido mais amplo que engloba inclusive os aracnídeos (carrapatos e ácaros). Filo Arthropoda O filo arthropoda
Entomologia Entomologia veterinária: é o estudo de insetos de importância veterinária, num sentido mais amplo que engloba inclusive os aracnídeos (carrapatos e ácaros). Filo Arthropoda O filo arthropoda
Boethus catarinae sp. nov.
 Acta Biol. Par., Curitiba, 31 (1, 2, 3, 4): 51-58. 2002 51 Boethus catarinae sp. nov. do Brasil (Hymenoptera, Ichneumonidae, Tryphoninae) 1 Boethus catarinae sp. nov. from Brazil (Hymenoptera, Ichneumonidae,
Acta Biol. Par., Curitiba, 31 (1, 2, 3, 4): 51-58. 2002 51 Boethus catarinae sp. nov. do Brasil (Hymenoptera, Ichneumonidae, Tryphoninae) 1 Boethus catarinae sp. nov. from Brazil (Hymenoptera, Ichneumonidae,
Universidade de São Paulo Instituto de Física de São Carlos Licenciatura em Ciências Exatas Biologia
 Universidade de São Paulo Instituto de Física de São Carlos Licenciatura em Ciências Exatas Biologia 2 2016 Aula Prática 7: ARTHROPODA UNIRAMIA: MYRIAPODA E INSECTA MYRIAPODA Os miriápodos possuem cabeça
Universidade de São Paulo Instituto de Física de São Carlos Licenciatura em Ciências Exatas Biologia 2 2016 Aula Prática 7: ARTHROPODA UNIRAMIA: MYRIAPODA E INSECTA MYRIAPODA Os miriápodos possuem cabeça
Carlos Guilherme C. Mielke 2, Olaf H. H. Mielke 3 & Mirna M. Casagrande 3
 Estudo comparado ado da morfologia ologia externa de Zaretis itys itylus (Westw estwood) e Agrias claudina annetta (Gray) (Lepidoptera, Nymphalidae, Charaxinae). axinae). III. Abdome 1 Carlos Guilherme
Estudo comparado ado da morfologia ologia externa de Zaretis itys itylus (Westw estwood) e Agrias claudina annetta (Gray) (Lepidoptera, Nymphalidae, Charaxinae). axinae). III. Abdome 1 Carlos Guilherme
Morfologia externa do adulto de Hemiargus hanno (Lepidoptera, Lycaenidae, Polyommatinae, Polyommatini). II. Região cervical, tórax e abdome
. II. Região cervical, tórax e abdome") 194 Morfologia externa do adulto de Hemiargus hanno (Lepidoptera, Lycaenidae, Polyommatinae, Polyommatini). II. Região cervical, tórax e abdome Marcelo Duarte Museu de Zoologia, Universidade de São Paulo,
194 Morfologia externa do adulto de Hemiargus hanno (Lepidoptera, Lycaenidae, Polyommatinae, Polyommatini). II. Região cervical, tórax e abdome Marcelo Duarte Museu de Zoologia, Universidade de São Paulo,
INSETOS-PRAGA NO BRASIL: LAGARTA-PRETA
 INSETOS-PRAGA NO BRASIL: LAGARTA-PRETA BOAS PRÁTICAS AGRÍCOLAS Soluções para um Mundo em Crescimento ÍNDICE Contexto Ocorrência Abrangência nacional Características Curiosidades Alerta 01 02 03 05 09 10
INSETOS-PRAGA NO BRASIL: LAGARTA-PRETA BOAS PRÁTICAS AGRÍCOLAS Soluções para um Mundo em Crescimento ÍNDICE Contexto Ocorrência Abrangência nacional Características Curiosidades Alerta 01 02 03 05 09 10
UNIVERSIDADE FEDERAL DO PARANÁ MANOELA KOWALCZUCK
 UNIVERSIDADE FEDERAL DO PARANÁ MANOELA KOWALCZUCK Grapholita motrix Berg, 1891(LEPIDOPTERA, TORTRICIDAE, OLETHREUTINAE, GRAPHOLITINI). IDENTIDADE TAXONÔMICA, MORFOLOGIA, ASPECTOS BIOLÓGICOS E RELAÇÕES
UNIVERSIDADE FEDERAL DO PARANÁ MANOELA KOWALCZUCK Grapholita motrix Berg, 1891(LEPIDOPTERA, TORTRICIDAE, OLETHREUTINAE, GRAPHOLITINI). IDENTIDADE TAXONÔMICA, MORFOLOGIA, ASPECTOS BIOLÓGICOS E RELAÇÕES
Espécies novas de Eucerinae Neotropicais e notas sobre Dasyhalonia phaeoptera Moure & Michener (Hymenoptera, Anthophoridae) 1
 1") Espécies novas de Eucerinae Neotropicais e notas sobre Dasyhalonia phaeoptera Moure & Michener (Hymenoptera, Anthophoridae) 1 Danúncia Urban 2 ABSTRACT. New species of neotropical Eucerinae and notes on
Espécies novas de Eucerinae Neotropicais e notas sobre Dasyhalonia phaeoptera Moure & Michener (Hymenoptera, Anthophoridae) 1 Danúncia Urban 2 ABSTRACT. New species of neotropical Eucerinae and notes on
ESTÁGIOS IMATUROS DE LOXOLOMIA JOHNSONI SCHAUS (LEPIDOPTERA, SATURNIIDAE, ARSENURINAE)
") ESTÁGIOS IMATUROS DE LOXOLOMIA JOHNSONI SCHAUS (LEPIDOPTERA, SATURNIIDAE, ARSENURINAE) Eurides Furtado 1 ABSTRACT. IMMATURE STAGES OF LOXOLOMIA.I0liNSONI SCHAUS (LEPIDOPTERA, SA. TURNIIDA E, ARSENURINAE).
ESTÁGIOS IMATUROS DE LOXOLOMIA JOHNSONI SCHAUS (LEPIDOPTERA, SATURNIIDAE, ARSENURINAE) Eurides Furtado 1 ABSTRACT. IMMATURE STAGES OF LOXOLOMIA.I0liNSONI SCHAUS (LEPIDOPTERA, SA. TURNIIDA E, ARSENURINAE).
Descrição dos Imaturos de Loxa deducta Walker e Pallantia macunaima Grazia (Heteroptera: Pentatomidae) em Ligustro, Ligustrum lucidum Ait.
 em Ligustro, Ligustrum lucidum Ait.") Março, 2001 Neotropical Entomology 30(1) 73 SISTEMÁTICA, MORFOLOGIA E FISIOLOGIA Descrição dos Imaturos de Loxa deducta Walker e Pallantia macunaima Grazia (Heteroptera: Pentatomidae) em Ligustro, Ligustrum
Março, 2001 Neotropical Entomology 30(1) 73 SISTEMÁTICA, MORFOLOGIA E FISIOLOGIA Descrição dos Imaturos de Loxa deducta Walker e Pallantia macunaima Grazia (Heteroptera: Pentatomidae) em Ligustro, Ligustrum
MORFOLOGIA EXTERNA DA OPERÁRIA DE Lestrimelitta ehrhardti (FRIESE, 1931) (HYMENOPTERA: MELIPONINI).
 (HYMENOPTERA: MELIPONINI).") Acta biol. Colomb., Vol. 15 N.º 2, 2010 131-148 MORFOLOGIA EXTERNA DA OPERÁRIA DE Lestrimelitta ehrhardti (FRIESE, 1931) (HYMENOPTERA: MELIPONINI). Worker External Morphology of Lestrimelitta ehrhardti
Acta biol. Colomb., Vol. 15 N.º 2, 2010 131-148 MORFOLOGIA EXTERNA DA OPERÁRIA DE Lestrimelitta ehrhardti (FRIESE, 1931) (HYMENOPTERA: MELIPONINI). Worker External Morphology of Lestrimelitta ehrhardti
DENISE QUIRINO TEODORO
 DENISE QUIRINO TEODORO CONTRIBUIÇÃO A O ESTUDO DA MORFOLOÇIA DO ADULTO DE PREPONA LAERTES (HÜBNER) (LEPIDOPTERA - NYM PHAUDAE - CHARAXINAE) Monografia apresentada ao Curso de Ciências Biológicas para a
DENISE QUIRINO TEODORO CONTRIBUIÇÃO A O ESTUDO DA MORFOLOÇIA DO ADULTO DE PREPONA LAERTES (HÜBNER) (LEPIDOPTERA - NYM PHAUDAE - CHARAXINAE) Monografia apresentada ao Curso de Ciências Biológicas para a
Características gerais
 Artrópodes Quem são? Muitas vezes, não percebemos a presença daqueles animais com corpos de formas estranhas e cores variadas, que vivem ao nosso redor, voam sobre nossas cabeças ou aqueles que se locomovem
Artrópodes Quem são? Muitas vezes, não percebemos a presença daqueles animais com corpos de formas estranhas e cores variadas, que vivem ao nosso redor, voam sobre nossas cabeças ou aqueles que se locomovem
Estudo Morfológico de Batrachedra nuciferae Hodges (Lepidotera: Coleophoridae)
") March - April 2004 173 SYSTEMATICS, MORPHOLOGY AND PHYSIOLOGY Estudo Morfológico de Batrachedra nuciferae Hodges (Lepidotera: Coleophoridae) SAÚL SÁNCHEZ-SOTO 1 E OCTAVIO NAKANO 2 1 Campus Tabasco, Colegio
March - April 2004 173 SYSTEMATICS, MORPHOLOGY AND PHYSIOLOGY Estudo Morfológico de Batrachedra nuciferae Hodges (Lepidotera: Coleophoridae) SAÚL SÁNCHEZ-SOTO 1 E OCTAVIO NAKANO 2 1 Campus Tabasco, Colegio
Percevejos predadores: o estado taxonômico e sistemático atual e perspectivas (Heteroptera: Pentatomidae: Asopinae)
") Percevejos predadores: o estado taxonômico e sistemático atual e perspectivas (Heteroptera: Pentatomidae: Asopinae) Talita Roell tali.roell@gmail.com Heteroptera Mais de 40mil espécies descritas Pentatomidae
Percevejos predadores: o estado taxonômico e sistemático atual e perspectivas (Heteroptera: Pentatomidae: Asopinae) Talita Roell tali.roell@gmail.com Heteroptera Mais de 40mil espécies descritas Pentatomidae
Morfologia dos estágios imaturos de Eupalea reinhardti Crotch (Coleoptera, Coccinellidae) e alguns dados biológicos 1
 e alguns dados biológicos 1") Morfologia dos estágios imaturos de Eupalea reinhardti Crotch (Coleoptera, Coccinellidae) e alguns dados biológicos 1 Flávia de Albuquerque Seródio Ferreira 2 Lúcia Massutti de Almeida 2 ABSTRACT. Morphology
Morfologia dos estágios imaturos de Eupalea reinhardti Crotch (Coleoptera, Coccinellidae) e alguns dados biológicos 1 Flávia de Albuquerque Seródio Ferreira 2 Lúcia Massutti de Almeida 2 ABSTRACT. Morphology
Notas complementares sobre a biologia de Rhescyntis pseudomartii Lemaire, 1980 (Lepidoptera, Adelocephalidae)1 ABSTRACT
1 ABSTRACT") REVISTA BRASILEIRA DE ZOOLOGIA Revta bras. Zool., 6 (4) : 6 4 9-6 5 7 31/XII/89 Notas complementares sobre a biologia de Rhescyntis pseudomartii Lemaire, 1980 (Lepidoptera, Adelocephalidae)1 Danúncia Urban
REVISTA BRASILEIRA DE ZOOLOGIA Revta bras. Zool., 6 (4) : 6 4 9-6 5 7 31/XII/89 Notas complementares sobre a biologia de Rhescyntis pseudomartii Lemaire, 1980 (Lepidoptera, Adelocephalidae)1 Danúncia Urban
Subfilo Uniramia. Superclasse Myriapoda Superclasse Hexapoda. Subfilo Myriapoda Subfilo Hexapoda
 MYRIAPODA Subfilo Uniramia Superclasse Myriapoda Superclasse Hexapoda ou Subfilo Myriapoda Subfilo Hexapoda Myriapoda (Gr. myria = inúmeros; podos = pés) - Lacraias, centopéias, piolhos- de- cobra, embuás,
MYRIAPODA Subfilo Uniramia Superclasse Myriapoda Superclasse Hexapoda ou Subfilo Myriapoda Subfilo Hexapoda Myriapoda (Gr. myria = inúmeros; podos = pés) - Lacraias, centopéias, piolhos- de- cobra, embuás,
Espécies novas de Ischnoptera (Blattellidae, Blattellinae) do Estado do Mato Grosso, Brasil e considerações sobre I. similis
 do Estado do Mato Grosso, Brasil e considerações sobre I. similis") 172 Espécies novas de Ischnoptera (Blattellidae, Blattellinae) do Estado do Mato Grosso, Brasil e considerações sobre I. similis Sonia Maria Lopes Departamento de Entomologia, Museu Nacional, Universidade
172 Espécies novas de Ischnoptera (Blattellidae, Blattellinae) do Estado do Mato Grosso, Brasil e considerações sobre I. similis Sonia Maria Lopes Departamento de Entomologia, Museu Nacional, Universidade
Ordem Trichoptera Kirby 1813 (Arthropoda: Insecta)
") Ordem Trichoptera Kirby 1813 (Arthropoda: Insecta) Adolfo Ricardo Calor Laboratório de Entomologia Aquática, FFCLRP, USP acalor@usp.br Introdução Trichoptera Kirby, 1813 compreendem a maior ordem de insetos
Ordem Trichoptera Kirby 1813 (Arthropoda: Insecta) Adolfo Ricardo Calor Laboratório de Entomologia Aquática, FFCLRP, USP acalor@usp.br Introdução Trichoptera Kirby, 1813 compreendem a maior ordem de insetos
SISTEMÁTICA DE Rhabdepyris KIEFFER (HYMENOPTERA, BETHYLIDAE) AMAZÔNICOS COM ANTENAS PECTINADAS
 AMAZÔNICOS COM ANTENAS PECTINADAS") SISTEMÁTICA DE Rhabdepyris KIEFFER (HYMENOPTERA, BETHYLIDAE) AMAZÔNICOS COM ANTENAS PECTINADAS Adriano da S. MAGESKY 1, Celso Oliveira AZEVEDO 1 RESUMO - São descritas e ilustradas Rhabdepyris opistolatus
SISTEMÁTICA DE Rhabdepyris KIEFFER (HYMENOPTERA, BETHYLIDAE) AMAZÔNICOS COM ANTENAS PECTINADAS Adriano da S. MAGESKY 1, Celso Oliveira AZEVEDO 1 RESUMO - São descritas e ilustradas Rhabdepyris opistolatus
1. Três: cabeça, tórax e abdômen (insetos) 2. Dois: cefalotórax e abdômen (aracnídeos)
 2. Dois: cefalotórax e abdômen (aracnídeos)") Silvestre 1. Metâmeros bem distintos na fase larvária. 2. Nos adultos se encontram fundidos. 3. Os metâmeros podem se apresentar agrupados em três regiões distintas (insetos) ou em duas (aracnídeos). 1.
Silvestre 1. Metâmeros bem distintos na fase larvária. 2. Nos adultos se encontram fundidos. 3. Os metâmeros podem se apresentar agrupados em três regiões distintas (insetos) ou em duas (aracnídeos). 1.
REVISTA BRASILEIRA DE ZOOLOGIA
 455 REVISTA BRASILEIRA DE ZOOLOGIA Revta bras. Zool., 5(3) 455-464 15.IX.1988 ESTÁGIOS IMATUROS DE SABULODES CABERATA CABERATA GUENéE, 1857 E SABULODES EXHONORATA GUENÉE, 1857 (LEPIDOPTERA, GEOMETRIDAE,
455 REVISTA BRASILEIRA DE ZOOLOGIA Revta bras. Zool., 5(3) 455-464 15.IX.1988 ESTÁGIOS IMATUROS DE SABULODES CABERATA CABERATA GUENéE, 1857 E SABULODES EXHONORATA GUENÉE, 1857 (LEPIDOPTERA, GEOMETRIDAE,
Fioravante Giacomel 2
 MORFOLOGIA E COMPORTAMENTO DAS FORMAS IMATURAS DE Psy/otoxus me/anospidus GIACOMEL (COLEOPTERA, CERAMBYCIDAE, LAMIINAE) 1 Fioravante Giacomel 2 ABSfRACf. MORPHOLOGY AND BEHAVIOR OF THE IMMATURE FORMS OF
MORFOLOGIA E COMPORTAMENTO DAS FORMAS IMATURAS DE Psy/otoxus me/anospidus GIACOMEL (COLEOPTERA, CERAMBYCIDAE, LAMIINAE) 1 Fioravante Giacomel 2 ABSfRACf. MORPHOLOGY AND BEHAVIOR OF THE IMMATURE FORMS OF
CAPÍTULO XVII. Ordem ZORAPTERA 117
 CAPÍTULO XVII Ordem ZORAPTERA 117 177. Caracteres. - Insetos pequenos (2 a 3 mm. de comprimento), com indivíduos, de ambos os sexos, apteros e alados, estes de 4 asas desiguais, as anteriores maiores que
CAPÍTULO XVII Ordem ZORAPTERA 117 177. Caracteres. - Insetos pequenos (2 a 3 mm. de comprimento), com indivíduos, de ambos os sexos, apteros e alados, estes de 4 asas desiguais, as anteriores maiores que
TÓRAX. Prof.: Gustavo Martins Pires
 TÓRAX Prof.: Gustavo Martins Pires INTRODUÇÃO O tórax é a porção mais superior do tronco e abriga órgãos fundamentais para a respiração, os pulmões, e para a circulação, o coração. Além disto, é atravessado
TÓRAX Prof.: Gustavo Martins Pires INTRODUÇÃO O tórax é a porção mais superior do tronco e abriga órgãos fundamentais para a respiração, os pulmões, e para a circulação, o coração. Além disto, é atravessado
Acta Biol. Par., Curitiba, 41 (3-4): KETI MARIA ROCHA ZANOL 2 Estrianna gen. nov. (Espécie tipo Estrianna sinopia sp. n.
: KETI MARIA ROCHA ZANOL 2 Estrianna gen. nov. (Espécie tipo Estrianna sinopia sp. n.") Acta Biol. Par., Curitiba, 41 (3-4): 67-73. 2012. 67 Estrianna gen. nov. (Hemiptera, Cicadellidae, Neobaliinae) e descrições de três novas espécies 1 Estrianna gen. nov. (Hemiptera, Cicadellidae, Neobaliinae)
Acta Biol. Par., Curitiba, 41 (3-4): 67-73. 2012. 67 Estrianna gen. nov. (Hemiptera, Cicadellidae, Neobaliinae) e descrições de três novas espécies 1 Estrianna gen. nov. (Hemiptera, Cicadellidae, Neobaliinae)
Reprodução e desenvolvimento. Prof. Jucelaine Haas
 Reprodução e desenvolvimento Prof. Jucelaine Haas Reprodução oviparidade (local depende do hábito) viviparidade (Glossina, tsé tsé) partenogênese (telítoca, arrenótoca, anfítoca Desenvolvimento pós-embrionário
Reprodução e desenvolvimento Prof. Jucelaine Haas Reprodução oviparidade (local depende do hábito) viviparidade (Glossina, tsé tsé) partenogênese (telítoca, arrenótoca, anfítoca Desenvolvimento pós-embrionário
UNIVERSIDADE ESTADUAL PAULISTA FACULDADE DE CIÊNCIAS AGRÁRIAS E VETERINÁRIAS DEPARTAMENTO DE FITOSSANIDADE Disciplina de Entomologia Básica
 UNIVERSIDADE ESTADUAL PAULISTA FACULDADE DE CIÊNCIAS AGRÁRIAS E VETERINÁRIAS DEPARTAMENTO DE FITOSSANIDADE Disciplina de Entomologia Básica AULA: CARACTERÍSTICAS DAS PRINCIPAIS ORDENS DE INSETOS DE IMPORTÂNCIA
UNIVERSIDADE ESTADUAL PAULISTA FACULDADE DE CIÊNCIAS AGRÁRIAS E VETERINÁRIAS DEPARTAMENTO DE FITOSSANIDADE Disciplina de Entomologia Básica AULA: CARACTERÍSTICAS DAS PRINCIPAIS ORDENS DE INSETOS DE IMPORTÂNCIA
A principal característica que diferencia os artrópodes dos demais invertebrados são as patas articuladas.
 Características A principal característica que diferencia os artrópodes dos demais invertebrados são as patas articuladas. A expressão patas articuladas vem do grego: artro, que significa "articulação",
Características A principal característica que diferencia os artrópodes dos demais invertebrados são as patas articuladas. A expressão patas articuladas vem do grego: artro, que significa "articulação",
Crustáceos (crusta= crosta ou pele grossa)
") Grupo Mandibulata Subfilos: Miriápodes: quilópodes (centopéias e lacraias) e diplópodes (piolho de cobra). Crustáceos Insetos Novidades evolutivas: Mandíbulas (importantes para triturar alimentos); Olhos
Grupo Mandibulata Subfilos: Miriápodes: quilópodes (centopéias e lacraias) e diplópodes (piolho de cobra). Crustáceos Insetos Novidades evolutivas: Mandíbulas (importantes para triturar alimentos); Olhos
INSETOS-PRAGA NO BRASIL: LAGARTA-DAS-VAGENS
 INSETOS-PRAGA NO BRASIL: LAGARTA-DAS-VAGENS BOAS PRÁTICAS AGRÍCOLAS Soluções para um Mundo em Crescimento ÍNDICE Contexto 01 Ocorrência 02 Abrangência nacional 03 Características 05 Curiosidades 09 Alerta
INSETOS-PRAGA NO BRASIL: LAGARTA-DAS-VAGENS BOAS PRÁTICAS AGRÍCOLAS Soluções para um Mundo em Crescimento ÍNDICE Contexto 01 Ocorrência 02 Abrangência nacional 03 Características 05 Curiosidades 09 Alerta
Morfologia externa dos estágios imaturos de heliconíneos neotropicais: I. Eueides isabella dianasa (Hübner, 1806) 1
 1") Revista Brasileira de Entomologia 46(4): 601-610 31.XII.2002 Morfologia externa dos estágios imaturos de heliconíneos neotropicais: I. Eueides isabella dianasa (Hübner, 1806) 1 Fabiano F. Antunes 2 Ayres
Revista Brasileira de Entomologia 46(4): 601-610 31.XII.2002 Morfologia externa dos estágios imaturos de heliconíneos neotropicais: I. Eueides isabella dianasa (Hübner, 1806) 1 Fabiano F. Antunes 2 Ayres
ALGUMAS CONSIDERAÇÕES BIOLÓGICAS EM SPODOPTERA FRUGIPERDA EM MILHO NO MUNICÍPIO DE ITUIUTABA (MG)
") ALGUMAS CONSIDERAÇÕES BIOLÓGICAS EM SPODOPTERA FRUGIPERDA EM MILHO NO MUNICÍPIO DE ITUIUTABA (MG) VILARINHO, E.C.1; ASSIS, J.M.F.1; CRUZ, I.2 1Dep. Ciências Agrárias. ISEPI/UEMG, Caixa Postal 431-38.300-000.
ALGUMAS CONSIDERAÇÕES BIOLÓGICAS EM SPODOPTERA FRUGIPERDA EM MILHO NO MUNICÍPIO DE ITUIUTABA (MG) VILARINHO, E.C.1; ASSIS, J.M.F.1; CRUZ, I.2 1Dep. Ciências Agrárias. ISEPI/UEMG, Caixa Postal 431-38.300-000.
15/03/2016 ESQUELETO APENDICULAR OSTEOLOGIA DO ESQUELETO APENDICULAR MEMBRO TORÁCICO. Constituído por ossos dos membros torácico e pélvico
 ESQUELETO APENDICULAR Constituído por ossos dos membros torácico e pélvico OSTEOLOGIA DO ESQUELETO APENDICULAR Ombro - Escápula Braço - Úmero Antebraço Rádio e Ulna Joelho - Carpos Canela - Metacarpos
ESQUELETO APENDICULAR Constituído por ossos dos membros torácico e pélvico OSTEOLOGIA DO ESQUELETO APENDICULAR Ombro - Escápula Braço - Úmero Antebraço Rádio e Ulna Joelho - Carpos Canela - Metacarpos
3/3/2017 FACULDADE EDUCACIONAL DE MEDIANEIRA. Principais funções da cabeça dos insetos: Olhos compostos e ocelos. Antenas.
 FACULDADE EDUCACIONAL DE MEDIANEIRA MORFOLOGIA DOS INSETOS Profª. Dra. Patrícia Bellon. Março/2017 MORFOLOGIA: CABEÇA DOS INSETOS Principais funções da cabeça dos insetos: MORFOLOGIA: CABEÇA DOS INSETOS
FACULDADE EDUCACIONAL DE MEDIANEIRA MORFOLOGIA DOS INSETOS Profª. Dra. Patrícia Bellon. Março/2017 MORFOLOGIA: CABEÇA DOS INSETOS Principais funções da cabeça dos insetos: MORFOLOGIA: CABEÇA DOS INSETOS
Acta Biol. Par., Curitiba, 46 (1-2):
:") Acta Biol. Par., Curitiba, 46 (1-2): 1-8. 2017. 1, Scaptotrigona guimaraesensis, uma nova espécie de abelha sem ferrão (Hym., Anthophila) do centro-oeste brasileiro Scaptotrigona guimaraesensis, a new
Acta Biol. Par., Curitiba, 46 (1-2): 1-8. 2017. 1, Scaptotrigona guimaraesensis, uma nova espécie de abelha sem ferrão (Hym., Anthophila) do centro-oeste brasileiro Scaptotrigona guimaraesensis, a new
Duas novas espécies de Mauesia (Cerambycidae, Lamiinae, Mauesini) e chave para identificação das espécies do gênero
 e chave para identificação das espécies do gênero") Volume 49(37):497 501, 2009 Duas novas espécies de Mauesia (Cerambycidae, Lamiinae, Mauesini) e chave para identificação das espécies do gênero Eleandro Moysés 1,2 Maria Helena M. Galileo 1,3 Abstract
Volume 49(37):497 501, 2009 Duas novas espécies de Mauesia (Cerambycidae, Lamiinae, Mauesini) e chave para identificação das espécies do gênero Eleandro Moysés 1,2 Maria Helena M. Galileo 1,3 Abstract
(Megalo = grande; pteron = asas) Carlos Augusto Silva de Azevêdo 1, Neusa Hamada 2
 Carlos Augusto Silva de Azevêdo 1, Neusa Hamada 2") 19 Ordem Megaloptera (Megalo = grande; pteron = asas) Carlos ugusto Silva de zevêdo 1, Neusa Hamada 2 Diagnose. Insetos neópteros de tamanho médio a grande (10-90 mm). Larva com o corpo deprimido dorso
19 Ordem Megaloptera (Megalo = grande; pteron = asas) Carlos ugusto Silva de zevêdo 1, Neusa Hamada 2 Diagnose. Insetos neópteros de tamanho médio a grande (10-90 mm). Larva com o corpo deprimido dorso
Espécies novas e notas sobre Nananthidium Moure (Hymenoptera, Apidae, Megachilinae) 1
 1") (Hymenoptera, Apidae, Megachilinae) 1 Espécies novas e notas sobre Nananthidium Moure 285 Danúncia Urban 2 1 Contribuição n o 1662 do Departamento de Zoologia, Universidade Federal do Paraná. 2 Departamento
(Hymenoptera, Apidae, Megachilinae) 1 Espécies novas e notas sobre Nananthidium Moure 285 Danúncia Urban 2 1 Contribuição n o 1662 do Departamento de Zoologia, Universidade Federal do Paraná. 2 Departamento
Acta Biol. Par., Curitiba, 32 (1, 2, 3, 4): DANÚNCIA URBAN 2
: DANÚNCIA URBAN 2") Acta Biol. Par., Curitiba, 32 (1, 2, 3, 4): 95-106. 2003. 95 Novos Dianthidiini neotropicais (Hymenoptera, Megachilidae) 1 New neotropical Dianthidiini (Hymenoptera, Megachilidae) DANÚNCIA URBAN 2 São
Acta Biol. Par., Curitiba, 32 (1, 2, 3, 4): 95-106. 2003. 95 Novos Dianthidiini neotropicais (Hymenoptera, Megachilidae) 1 New neotropical Dianthidiini (Hymenoptera, Megachilidae) DANÚNCIA URBAN 2 São
Revista Árvore ISSN: Universidade Federal de Viçosa Brasil
 Revista Árvore ISSN: 0100-6762 r.arvore@ufv.br Universidade Federal de Viçosa Brasil Fagundes Pereira, Fabricio; Cola Zanuncio, José; Hendrik Mielke, Olaf Hermann; de Oliveira, Juliana Maria; Costa Paiva,
Revista Árvore ISSN: 0100-6762 r.arvore@ufv.br Universidade Federal de Viçosa Brasil Fagundes Pereira, Fabricio; Cola Zanuncio, José; Hendrik Mielke, Olaf Hermann; de Oliveira, Juliana Maria; Costa Paiva,
Morfología externa dos adultos de Phthia picta (Drury, 1770) (Hemiptera, Coreidae)
 (Hemiptera, Coreidae)") Bol. San. Veg. Plagas, 29: 249-253, 2003 Morfología externa dos adultos de Phthia picta (Drury, 1770) (Hemiptera, Coreidae) R. A. DA SILVA, G. S. CARVALHO, P. S. FLORES O objetivo deste trabalho foi descrever
Bol. San. Veg. Plagas, 29: 249-253, 2003 Morfología externa dos adultos de Phthia picta (Drury, 1770) (Hemiptera, Coreidae) R. A. DA SILVA, G. S. CARVALHO, P. S. FLORES O objetivo deste trabalho foi descrever
Artrópodes. Filo Arthropoda (arthron = articulações e podos = pés)
") Artrópodes Filo Arthropoda (arthron = articulações e podos = pés) Características gerais Mais numeroso do planeta (1.000.000 espécies) Exemplos: insetos, aranhas, caranguejos Vivem em diversos ambientes
Artrópodes Filo Arthropoda (arthron = articulações e podos = pés) Características gerais Mais numeroso do planeta (1.000.000 espécies) Exemplos: insetos, aranhas, caranguejos Vivem em diversos ambientes
NOTAS SÔBRE Pholus vitis (L., 1758) (Lep., Sphingidae)
 (Lep., Sphingidae)") NOTAS SÔBRE Pholus vitis (L., 1758) (Lep., Sphingidae) LUIZ GONZAGA E. LORDELLO * (Assistente da Cadeira de Zoologia da Escola Superior de Agricultura "Luiz de Queiroz" da Universidade de São Paulo) A
NOTAS SÔBRE Pholus vitis (L., 1758) (Lep., Sphingidae) LUIZ GONZAGA E. LORDELLO * (Assistente da Cadeira de Zoologia da Escola Superior de Agricultura "Luiz de Queiroz" da Universidade de São Paulo) A
VANESSA ALVES NORONHA. ESTUDO DA MORFOLOGIA DE Gypona (Marganalana) hiata DELONG & FREYTAG, 1964 (HEMIPTERA, AUCHENORRHYNCHA, GYPONINAE)
 hiata DELONG & FREYTAG, 1964 (HEMIPTERA, AUCHENORRHYNCHA, GYPONINAE)") VANESSA ALVES NORONHA ESTUDO DA MORFOLOGIA DE Gypona (Marganalana) hiata DELONG & FREYTAG, 1964 (HEMIPTERA, AUCHENORRHYNCHA, GYPONINAE) CURITIBA 2000 VANESSA ALVES NORONHA ESTUDO DA MORFOLOGIA DE Gypona
VANESSA ALVES NORONHA ESTUDO DA MORFOLOGIA DE Gypona (Marganalana) hiata DELONG & FREYTAG, 1964 (HEMIPTERA, AUCHENORRHYNCHA, GYPONINAE) CURITIBA 2000 VANESSA ALVES NORONHA ESTUDO DA MORFOLOGIA DE Gypona
ande (Lepidoptera, urinae
 Morfologia ologia externa do adulto de Almeidaia aidae Mielke e & Casagrande ande (Lepidoptera, Saturniidae, Arsen senur urinae, Almeidaiini). III. Abdome 1 Amabílio J. A. de Camargo 2 ; Mirna M. Casagrande
Morfologia ologia externa do adulto de Almeidaia aidae Mielke e & Casagrande ande (Lepidoptera, Saturniidae, Arsen senur urinae, Almeidaiini). III. Abdome 1 Amabílio J. A. de Camargo 2 ; Mirna M. Casagrande
Revta bras. Ent. 32(2): OPERA OPILIOLOGICA VARIA. XXI. (OPILIONES, GONYLEPTIDAE) ABSTRACT
: OPERA OPILIOLOGICA VARIA. XXI. (OPILIONES, GONYLEPTIDAE) ABSTRACT") Revta bras. Ent. 32(2):233-237. 26.09.1988 8 OPERA OPILIOLOGICA VARIA. XXI. (OPILIONES, GONYLEPTIDAE) Helia E.M.Soares1 Benedicto A.M. Soares* ABSTRACT Stefanesia avramae, gen. n., sp.n., described from
Revta bras. Ent. 32(2):233-237. 26.09.1988 8 OPERA OPILIOLOGICA VARIA. XXI. (OPILIONES, GONYLEPTIDAE) Helia E.M.Soares1 Benedicto A.M. Soares* ABSTRACT Stefanesia avramae, gen. n., sp.n., described from
ESCOLA SUPERIOR DE AGRICULTURA LUIZ DE QUEIROZ - USP DEPARTAMENTO DE ENTOMOLOGIA E ACAROLOGIA
 ESCOLA SUPERIOR DE AGRICULTURA LUIZ DE QUEIROZ - USP DEPARTAMENTO DE ENTOMOLOGIA E ACAROLOGIA Disciplina: Blattaria-Isoptera Bibliografia Recomendada: Entomologia Agrícola p. 34-36 Ortopteroides Aparelho
ESCOLA SUPERIOR DE AGRICULTURA LUIZ DE QUEIROZ - USP DEPARTAMENTO DE ENTOMOLOGIA E ACAROLOGIA Disciplina: Blattaria-Isoptera Bibliografia Recomendada: Entomologia Agrícola p. 34-36 Ortopteroides Aparelho
PRIMEIRO REGISTRO DA FAMÍLIA SCELEMBIIDAE (EMBIOPTERA) PARA O BRASIL. Cristiano Machado Teixeira 1 Mikael Bolke Araújo 2 Flávio Roberto Mello Garcia 3
 PARA O BRASIL. Cristiano Machado Teixeira 1 Mikael Bolke Araújo 2 Flávio Roberto Mello Garcia 3") PRIMEIRO REGISTRO DA FAMÍLIA SCELEMBIIDAE (EMBIOPTERA) PARA O BRASIL Cristiano Machado Teixeira 1 Mikael Bolke Araújo 2 Flávio Roberto Mello Garcia 3 RESUMO: São registradas a primeira ocorrência da família
PRIMEIRO REGISTRO DA FAMÍLIA SCELEMBIIDAE (EMBIOPTERA) PARA O BRASIL Cristiano Machado Teixeira 1 Mikael Bolke Araújo 2 Flávio Roberto Mello Garcia 3 RESUMO: São registradas a primeira ocorrência da família
Uma nova espécie de Schizomyia (Diptera, Cecidomyiidae), indutora de galhas nos botões florais de Jacquemontia holosericea (Convolvulaceae)
, indutora de galhas nos botões florais de Jacquemontia holosericea (Convolvulaceae)") Uma nova espécie de Schizomyia (Diptera, Cecidomyiidae), indutora de galhas nos botões florais de Jacquemontia holosericea (Convolvulaceae) 356 Maia & Araújo Valéria Cid Maia 1 & Walter Santos de Araújo
Uma nova espécie de Schizomyia (Diptera, Cecidomyiidae), indutora de galhas nos botões florais de Jacquemontia holosericea (Convolvulaceae) 356 Maia & Araújo Valéria Cid Maia 1 & Walter Santos de Araújo
Dentre as diversas espécies de corós. Paulo Roberto V. S. Pereira1, José Roberto Salvadori1. Ministério da Agricultura, Pecuária e Abastecimento
 Ministério da Agricultura, Pecuária e Abastecimento 204 ISSN 1517-4964 Dezembro, 2006 Passo Fundo, RS Fotos/Imagens: Paulo Roberto V. S. Pereira Guia para identificação de corós rizófagos (Coleoptera:
Ministério da Agricultura, Pecuária e Abastecimento 204 ISSN 1517-4964 Dezembro, 2006 Passo Fundo, RS Fotos/Imagens: Paulo Roberto V. S. Pereira Guia para identificação de corós rizófagos (Coleoptera:
SHILAP Revista de Lepidopterología ISSN: Sociedad Hispano-Luso-Americana de Lepidopterología. España
 SHILAP Revista de Lepidopterología ISSN: 0300-5267 avives@eresmas.net Sociedad Hispano-Luso-Americana de Lepidopterología España Mielke, C. G. C.; Furtado, E. Quatro espécies novas do genus Periphora Hübner,
SHILAP Revista de Lepidopterología ISSN: 0300-5267 avives@eresmas.net Sociedad Hispano-Luso-Americana de Lepidopterología España Mielke, C. G. C.; Furtado, E. Quatro espécies novas do genus Periphora Hübner,
Deltocephalinae): primeiro registro no Brasil e descrição de duas espécies novas 1
: primeiro registro no Brasil e descrição de duas espécies novas 1") Bandaromimus Linna-vuori (Homoptera, Cicadellidae, Deltocephalinae): primeiro registro no Brasil e descrição de duas espécies novas 1 Keti Maria Rocha Zanol 2 ABSTRACT. 8andaromimus Linnavuori (Homoptera,
Bandaromimus Linna-vuori (Homoptera, Cicadellidae, Deltocephalinae): primeiro registro no Brasil e descrição de duas espécies novas 1 Keti Maria Rocha Zanol 2 ABSTRACT. 8andaromimus Linnavuori (Homoptera,
Imaturos de pentatomídeos (Hemiptera, Heteroptera): morfologia ABSTRACT
: morfologia ABSTRACT") Imaturos de pentatomídeos (Hemiptera, Heteroptera): morfologia... 81 IMATUROS DE PENTATOMÍDEOS (HEMIPTERA, HETEROPTERA): MORFOLOGIA E BIOLOGIA DE ACROSTERNUM OBSTINATUM 1 Viviana C. Matesco 2 Nora D. F.
Imaturos de pentatomídeos (Hemiptera, Heteroptera): morfologia... 81 IMATUROS DE PENTATOMÍDEOS (HEMIPTERA, HETEROPTERA): MORFOLOGIA E BIOLOGIA DE ACROSTERNUM OBSTINATUM 1 Viviana C. Matesco 2 Nora D. F.
NOVOS GÊNEROS E ESPÉCIES DE HOPLOPHORIONINI (HEMIPTERA, MEMBRACIDAE, MEMBRACINAE)
") NOVOS GÊNEROS E ESPÉCES DE HOPLOPHORONN (HEMPTERA, MEMBRACDAE, MEMBRACNAE) Antonio J. Creão-Duarle 1 ABSTRACT. NEW GENERA ANO NEW SPECES OF HOPLQPHORONlN (HEMPTERA, MEM BRACDAE, MEMBRACNAE). Two new genera
NOVOS GÊNEROS E ESPÉCES DE HOPLOPHORONN (HEMPTERA, MEMBRACDAE, MEMBRACNAE) Antonio J. Creão-Duarle 1 ABSTRACT. NEW GENERA ANO NEW SPECES OF HOPLQPHORONlN (HEMPTERA, MEM BRACDAE, MEMBRACNAE). Two new genera
Do grego di= dois e pteros= asas, sendo assim chamados devido a principal característica desta classe ser a presença de somente um par de asas
 ORDEM DIPTERA M A R C O S P A U L O B E Z M A U R I C I O S C A R A B E L O T M A T H E U S S O A R E S M I C A E L B R U N E L L I P A T R Í C I A L U M E R T Z Do grego di= dois e pteros= asas, sendo
ORDEM DIPTERA M A R C O S P A U L O B E Z M A U R I C I O S C A R A B E L O T M A T H E U S S O A R E S M I C A E L B R U N E L L I P A T R Í C I A L U M E R T Z Do grego di= dois e pteros= asas, sendo
ESTÁGIOS IMATUROS DE OEBALUS POECILUS (HETEROPTERA, PENTATOMIDAE) 1
 1") Estágios imaturos de Oebalus poecilus (Heteroptera, Pentatomidae) 89 ESTÁGIOS IMATUROS DE OEBALUS POECILUS (HETEROPTERA, PENTATOMIDAE) 1 ABSTRACT Caroline Greve 2 Nora D. F. Fortes 3 Jocélia Grazia 2 IMMATURES
Estágios imaturos de Oebalus poecilus (Heteroptera, Pentatomidae) 89 ESTÁGIOS IMATUROS DE OEBALUS POECILUS (HETEROPTERA, PENTATOMIDAE) 1 ABSTRACT Caroline Greve 2 Nora D. F. Fortes 3 Jocélia Grazia 2 IMMATURES
Artrópodes. Profº Fernando Belan - BIOLOGIA MAIS
 Artrópodes Profº Fernando Belan - BIOLOGIA MAIS Artrópodes Triblásticos Celomados Protostômios Simetria Bilateral Artrópodes Sistema nervosos ganglionar ventral hiponeuro. Sistema digestório completo Sistema
Artrópodes Profº Fernando Belan - BIOLOGIA MAIS Artrópodes Triblásticos Celomados Protostômios Simetria Bilateral Artrópodes Sistema nervosos ganglionar ventral hiponeuro. Sistema digestório completo Sistema
Occurrence of Fidicinoides pauliensis Boulard & Martinelli, 1996 (Hemiptera: Cicadidae) in coffee plant in Tapiratiba, SP
 in coffee plant in Tapiratiba, SP") From the SelectedWorks of Rodrigo Souza Santos RSS 2007 Occurrence of Fidicinoides pauliensis Boulard & Martinelli, 1996 (Hemiptera: Cicadidae) in coffee plant in Tapiratiba, SP Rodrigo Souza Santos, RSS
From the SelectedWorks of Rodrigo Souza Santos RSS 2007 Occurrence of Fidicinoides pauliensis Boulard & Martinelli, 1996 (Hemiptera: Cicadidae) in coffee plant in Tapiratiba, SP Rodrigo Souza Santos, RSS
Ordem Ephemeroptera (Arthropoda: Insecta)
") Ordem Ephemeroptera (Arthropoda: Insecta) Rodolfo Mariano Lopes da Silva Laboratório de Entomologia Aquática, FFCLRP, USP rodolfo@usp.br Introdução A ordem compreende insetos com adultos de hábitos terrestres
Ordem Ephemeroptera (Arthropoda: Insecta) Rodolfo Mariano Lopes da Silva Laboratório de Entomologia Aquática, FFCLRP, USP rodolfo@usp.br Introdução A ordem compreende insetos com adultos de hábitos terrestres
DESCRIÇÃO DE UMA NOVA ESPÉCIE DE HYMENOEPIMECIS VIERECK (HYMENOPTERA, ICHNEUMONIDAE, PIMPLINAE) PARASITÓIDE DE Leucauge Roseosignata
 PARASITÓIDE DE Leucauge Roseosignata") 77 DESCRIÇÃO DE UMA NOVA ESPÉCIE DE HYMENOEPIMECIS VIERECK (HYMENOPTERA, ICHNEUMONIDAE, PIMPLINAE) PARASITÓIDE DE Leucauge Roseosignata Sobczak J.F. 1. Loffredo. A.P.S. 1 Gonzaga M.O. 2 Pentado-Dias. A.M.M.
77 DESCRIÇÃO DE UMA NOVA ESPÉCIE DE HYMENOEPIMECIS VIERECK (HYMENOPTERA, ICHNEUMONIDAE, PIMPLINAE) PARASITÓIDE DE Leucauge Roseosignata Sobczak J.F. 1. Loffredo. A.P.S. 1 Gonzaga M.O. 2 Pentado-Dias. A.M.M.
DEIPARTAMENTO DE ZOOLOGIA. DOIS NOVOS CiÊNEROS E TRÊS NOVAS ESPÉCIES DE OPILIOES BRASILEIROS (*) POR INTRODUÇÁO
 POR INTRODUÇÁO") Vol. V, N." 26 - pp. 243-250 PAPÉIS AVULSOS DO DEIPARTAMENTO DE ZOOLOGIA SECRETARIA DA AGRICULTURA - S. PAULO - BRASIL DOIS NOVOS CiÊNEROS E TRÊS NOVAS ESPÉCIES DE OPILIOES BRASILEIROS (*) POR INTRODUÇÁO
Vol. V, N." 26 - pp. 243-250 PAPÉIS AVULSOS DO DEIPARTAMENTO DE ZOOLOGIA SECRETARIA DA AGRICULTURA - S. PAULO - BRASIL DOIS NOVOS CiÊNEROS E TRÊS NOVAS ESPÉCIES DE OPILIOES BRASILEIROS (*) POR INTRODUÇÁO
Morfologia externa. Rotraut A. G. B. Consoli Ricardo Lourenço de Oliveira
 Morfologia externa Rotraut A. G. B. Consoli Ricardo Lourenço de Oliveira SciELO Books / SciELO Livros / SciELO Libros CONSOLI, RAGB., and OLIVEIRA, RL. Principais mosquitos de importância sanitária no
Morfologia externa Rotraut A. G. B. Consoli Ricardo Lourenço de Oliveira SciELO Books / SciELO Livros / SciELO Libros CONSOLI, RAGB., and OLIVEIRA, RL. Principais mosquitos de importância sanitária no
Description of New Species of Fidicinoides Boulard & Martinelli (Hemiptera: Cicadidae) from Brazil
 from Brazil") From the SelectedWorks of Rodrigo Souza Santos RSS 2009 Description of New Species of Fidicinoides Boulard & Martinelli (Hemiptera: Cicadidae) from Brazil Rodrigo Souza Santos, RSS Available at: http://works.bepress.com/rodrigo_santos/7/
From the SelectedWorks of Rodrigo Souza Santos RSS 2009 Description of New Species of Fidicinoides Boulard & Martinelli (Hemiptera: Cicadidae) from Brazil Rodrigo Souza Santos, RSS Available at: http://works.bepress.com/rodrigo_santos/7/
Espécies novas de Blaptica do Rio Grande do Sul, Brasil (Blattaria, Blaberidae, Blaberinae)
") 93 Espécies novas de Blaptica do Rio Grande do Sul, Brasil (Blattaria, Blaberidae, Blaberinae) Sonia Maria Lopes 1 & Edivar Heeren de Oliveira 1 1. Depto de Entomologia, Museu Nacional, Universidade Federal
93 Espécies novas de Blaptica do Rio Grande do Sul, Brasil (Blattaria, Blaberidae, Blaberinae) Sonia Maria Lopes 1 & Edivar Heeren de Oliveira 1 1. Depto de Entomologia, Museu Nacional, Universidade Federal
ESTUDO DOS LEPIDÓPTEROS (BORBOLETAS) DA RESERVA ECOLÓGICA DO CAMPUS UEG, ANÁPOLIS GO. Graduandas do curso de Ciências Biológicas, UnuCET Anápolis UEG
 DA RESERVA ECOLÓGICA DO CAMPUS UEG, ANÁPOLIS GO. Graduandas do curso de Ciências Biológicas, UnuCET Anápolis UEG") ESTUDO DOS LEPIDÓPTEROS (BORBOLETAS) DA RESERVA ECOLÓGICA DO CAMPUS UEG, ANÁPOLIS GO Josiane Lima da Costa 1 ; Monique de Souza Paulino 1 ; Cynthia Aparecida Arossa Alves Soares 2 1 Graduandas do curso
ESTUDO DOS LEPIDÓPTEROS (BORBOLETAS) DA RESERVA ECOLÓGICA DO CAMPUS UEG, ANÁPOLIS GO Josiane Lima da Costa 1 ; Monique de Souza Paulino 1 ; Cynthia Aparecida Arossa Alves Soares 2 1 Graduandas do curso
Empresa Brasileira de Pesquisa Agropecuária Embrapa Amazônia Oriental Ministério da Agricultura, Pecuária e Abastecimento
 Empresa Brasileira de Pesquisa Agropecuária Embrapa Amazônia Oriental Ministério da Agricultura, Pecuária e Abastecimento Embrapa Belém, PA 2014 MONITORAMENTO DE Diaphorina citri EM CITROS NA EMBRAPA AMAZÔNIA
Empresa Brasileira de Pesquisa Agropecuária Embrapa Amazônia Oriental Ministério da Agricultura, Pecuária e Abastecimento Embrapa Belém, PA 2014 MONITORAMENTO DE Diaphorina citri EM CITROS NA EMBRAPA AMAZÔNIA
Aos meus pais Angelina e Rufino e minha irmã Valdirene.
 Aos meus pais Angelina e Rufino e minha irmã Valdirene. i AGRADECIMENTOS Ao professor Armando Carlos Cervi, pela orientação, ensinamentos, compreensão e disponibilidade. À professora Rosângela Capuano
Aos meus pais Angelina e Rufino e minha irmã Valdirene. i AGRADECIMENTOS Ao professor Armando Carlos Cervi, pela orientação, ensinamentos, compreensão e disponibilidade. À professora Rosângela Capuano
Designação de lectótipos de algumas espécies sul-americanas de Anthidium Fabricius (Hymenoptera, Megachilidae) 1
 1") Designação de lectótipos de algumas espécies sul-americanas de Anthidium Fabricius (Hymenoptera, Megachilidae) 1 Danúncia Urban 2 ABSTRACT. Designation of Lectotypes of some South American species of Anthidium
Designação de lectótipos de algumas espécies sul-americanas de Anthidium Fabricius (Hymenoptera, Megachilidae) 1 Danúncia Urban 2 ABSTRACT. Designation of Lectotypes of some South American species of Anthidium
CARACTERÍSTICAS MORFOLÓGICAS DE Rolepa unimoda (Dognin, 1923) (Lepidoptera, Lymantriidae)
 (Lepidoptera, Lymantriidae)") CARACTERÍSTICAS MORFOLÓGICAS DE Rolepa unimoda (Dognin, 1923) (Lepidoptera, Lymantriidae) Otávio Peres Filho* Evoneo Berti Filho** RESUMO Este trabalho relata a descrição morfológica de Rolepa unimoda
CARACTERÍSTICAS MORFOLÓGICAS DE Rolepa unimoda (Dognin, 1923) (Lepidoptera, Lymantriidae) Otávio Peres Filho* Evoneo Berti Filho** RESUMO Este trabalho relata a descrição morfológica de Rolepa unimoda
Galatheoidea Chirostylidae Uroptycbus uncifer (A. Milne Edwards, 1880)
") Anomura Galatheoidea Chirostylidae -- Uroptycbus uncifer (A. Milne Edwards, 1880) Diagnose : Esta espécie se diferencia essencialmente de U. nitidus pelas dimensões de seus quelípodos que são mais largos
Anomura Galatheoidea Chirostylidae -- Uroptycbus uncifer (A. Milne Edwards, 1880) Diagnose : Esta espécie se diferencia essencialmente de U. nitidus pelas dimensões de seus quelípodos que são mais largos
4ª Jornada Científica e Tecnológica e 1º Simpósio de Pós-Graduação do IFSULDEMINAS 16, 17 e 18 de outubro de 2012, Muzambinho MG
 4ª Jornada Científica e Tecnológica e 1º Simpósio de Pós-Graduação do IFSULDEMINAS 16, 17 e 18 de outubro de 2012, Muzambinho MG Diversidade de Insetos em Plantas Epífitas no Instituto Federal do Sul De
4ª Jornada Científica e Tecnológica e 1º Simpósio de Pós-Graduação do IFSULDEMINAS 16, 17 e 18 de outubro de 2012, Muzambinho MG Diversidade de Insetos em Plantas Epífitas no Instituto Federal do Sul De
Coronalidia, gênero novo de Neocoelidiinae (Hemiptera, Cicadellidae) do Equador 1
 do Equador 1") (Hemiptera, Cicadellidae) do Equador 1 294 Marques-Costa & Cavichioli Ana Paula Marques-Costa 2 & Rodney Ramiro Cavichioli 2 1 Contribuição número 1672 do Departamento de Zoologia, Universidade Federal
(Hemiptera, Cicadellidae) do Equador 1 294 Marques-Costa & Cavichioli Ana Paula Marques-Costa 2 & Rodney Ramiro Cavichioli 2 1 Contribuição número 1672 do Departamento de Zoologia, Universidade Federal
COLEÇÃO ENTOMOLÓGICA RESUMO
 COLEÇÃO ENTOMOLÓGICA Alexandre Felix da Silva Chiesa1 ; Guilherme Modena2 Marcelo Diel3 RESUMO A entomologia é a parte da biologia que estuda os insetos. Uma das melhores formas de estudar insetos é fazer
COLEÇÃO ENTOMOLÓGICA Alexandre Felix da Silva Chiesa1 ; Guilherme Modena2 Marcelo Diel3 RESUMO A entomologia é a parte da biologia que estuda os insetos. Uma das melhores formas de estudar insetos é fazer
Ordem Hemiptera Subordem Heteroptera. Nome comum: Percevejos
 Ordem Hemiptera Subordem Heteroptera Nome comum: Percevejos Identificação do hábito alimentar Fitófagos O rostro é reto, longo e delgado; apresentase subdividido em quatro segmentos; quando em repouso,
Ordem Hemiptera Subordem Heteroptera Nome comum: Percevejos Identificação do hábito alimentar Fitófagos O rostro é reto, longo e delgado; apresentase subdividido em quatro segmentos; quando em repouso,
DESCRIÇÃO DE DUAS ESPÉCIES NOVAS DE Elmohardyia RAFAEL (DIPTERA, PEPUNCULIDAE) DA REGIÃO AMAZÔNICA
 DA REGIÃO AMAZÔNICA") DESCRIÇÃO DE DUAS ESPÉCIES NOVAS DE Elmohardyia RAFAEL (DIPTERA, PEPUNCULIDAE) DA REGIÃO AMAZÔNICA Maria Denise da Silva MENEZES 1 José Albertino RAFAEL 1 2 RESUMO Duas espécies amazônicas de Elmohardyia
DESCRIÇÃO DE DUAS ESPÉCIES NOVAS DE Elmohardyia RAFAEL (DIPTERA, PEPUNCULIDAE) DA REGIÃO AMAZÔNICA Maria Denise da Silva MENEZES 1 José Albertino RAFAEL 1 2 RESUMO Duas espécies amazônicas de Elmohardyia